圆齿野鸦椿大小孢子发生及雌雄配子体形成
2021-09-29万瑞琪邵晓雪吴凌敏涂淑萍
万瑞琪,邵晓雪,吴凌敏,涂淑萍*
(1.江西农业大学 园林与艺术学院,江西 南昌 330045;2.江西省景德镇市林科所,江西 景德镇 333000;3.江西省景德镇市烟草专卖局,江西 景德镇 333000)
【研究意义】圆齿野鸦椿(Euscaphis konishiiHayata)为省沽油科野鸦椿属常绿小乔木,是我国特有的药、赏两用树种,野生资源主要分布在广东、海南、广西、福建、江西、湖南、贵州和四川等地[1]。圆齿野鸦椿结果量大,观果期长达7个月之久,跨秋、冬和翌年春三季,种子粘挂在反卷、形似蝴蝶的内果皮上,犹如满树红花上点缀着的黑珍珠,观赏价值极高,还可丰富秋冬季园林景观和生物多样性。目前,野鸦椿属植物中已发现的化学成分主要有三萜类、黄酮类和酚酸类等,具有抗肿瘤、抗肝纤维化和抗炎抑菌等药理活性[2-3]。因此,大力开发利用和推广种植该乡土树种,对于促进地方经济发展有着极其重要的意义。种苗培育是开发利用优良乡土植物资源的关键,目前,圆齿野鸦椿育苗仍以播种繁殖为主,种子质量是其发芽率和培育壮苗的关键。此外,播种繁殖在圆齿野鸦椿优良种源家系的选择和杂交育种方面亦具有重要地位。调查研究过程中发现,该树种存在空籽率高,种子发芽率低且具有深休眠现象,从而大大降低了其种苗繁育效能。【前人研究进展】目前国内学者已在野鸦椿属植物种质资源[4-5]、生态生物学特性[6]、种苗繁育[7-10]、种子休眠[11-14]等方面进行了大量研究。【本研究切入点】植物种子质量与其胚胎发育关系密切,然而关于圆齿野鸦椿生殖生物学及胚胎学方面的研究极为匮乏。【拟解决的关键问题】研究拟采用石蜡切片法观察圆齿野鸦椿大小孢子发生与雌雄配子体形成,旨在从胚胎学方面探讨其空籽率高、种子发芽率低的原因。
1 材料与方法
1.1 试验材料
试验材料取自江西农业大学风景园林实训基地(28°46'N,115°55'E,海拔50 m)种植的12 年生圆齿野鸦椿实生苗。
1.2 试验方法
1.2.1 圆齿野鸦椿大小孢子发生和雌雄配子体形成 于2015年5月起,接近开花时每天观察一次;开花期间,每隔几个小时观察一次,采集不同发育阶段的花芽,将其外部鳞片剥离后处理成小份,放入含70%FAA固定液中抽真空保存。石蜡切片法[15]制片,电子显微镜下观察拍照。
1.2.2 圆齿野鸦椿花粉形态观察 采集当天开放的新鲜花朵与其花药,用双面胶处理后喷金,在显微镜下观察拍照[15]。
2 结果与分析
2.1 圆齿野鸦椿小孢子发生与雄配子体形成
2.1.1 小孢子发生 雄蕊多为5 枚,每枚含4 个对称发育小孢子囊。花药发育初期,其横切面类似于梯形的结构(图1a)。随后其四周原表皮下第一层细胞分裂,形成体积较大、细胞核明显的孢原细胞(图1b)。孢原细胞分裂形成周缘细胞和初生造孢细胞。周缘细胞后形成花药壁,由内到外依次为绒毡层、中层、药室内壁和表皮(图1c);初生造孢细胞最终形成小孢子母细胞(图1d)。

图1 小孢子发生Fig.1 Microsporogenesis
2.1.2 雄配子体形成 小孢子母细胞经两次减数分裂形成4 个子细胞,由胼胝质相连(图2a)呈四面体排列。后期胼胝质分解,4 个子细胞分散逐步发育为雄配子体。小液泡逐渐发育,将细胞核挤向靠近细胞壁的位置,称为单核靠边期(图2b)。靠近细胞壁的小孢子有丝分裂形成两个大小不一的核(大的为营养核,小的为生殖核),细胞质也被分为大小不等的两部分(图2c)。绒毡层为小孢子发育提供营养,至花粉粒成熟时逐渐消失(图2d)。其成熟花粉为2-细胞型。
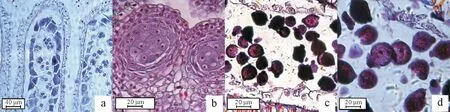
图2 雄配子体形成Fig.2 Formation of male gametophytes
2.1.3 花药壁发育 开始发育至花药成熟,其花药壁表皮细胞从方形变化至扁平,细胞排列从有序变化至不规则。随着花药壁发育,药室内壁仅有的一层细胞逐步延长加厚成纤维网状,以帮助花粉释放。中层细胞在减数分裂期间被逐渐吸收解体。绒毡层细胞起初为单核,细胞质浓,液泡少且小。减数分裂期间,绒毡层细胞分裂成二核至多核,呈不规则松散排列。当中层细胞被吸收分解,绒毡层亦分解,此时花粉成熟。部分空腔(图3a)或花粉败育(图3b),则是源于该药囊绒毡层解体过程异常。
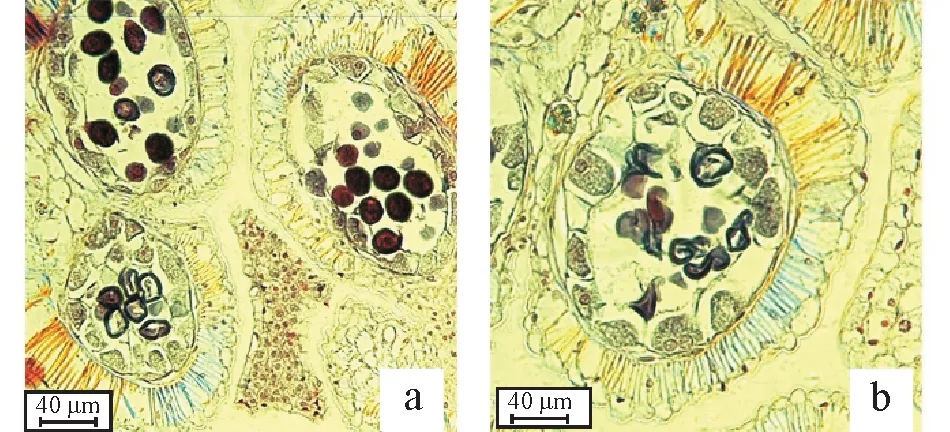
图3 花粉败育Fig.3 Pollen abortion
2.2 圆齿野鸦椿大孢子发生与雌配子体形成
2.2.1 大孢子发生 由图4 可知,圆齿野鸦椿属上位子房,3 室,每室3~4 枚横生胚珠(图4a),中轴胎座(图4b)。孢原细胞位于胚珠顶部表皮下方,其逐渐发育成核大、质浓的大孢子母细胞(图4c);后经两次减数分裂,形成4个呈线性排列的子细胞(图4d),有少部分呈“田”字形排列(图4e)。部分大孢子母细胞高度液泡化,不见核仁,未能进行减数分裂。靠近珠孔端的3个细胞退化,远离珠孔端的一个细胞发育成单核胚囊,称之为大孢子(图4f)。
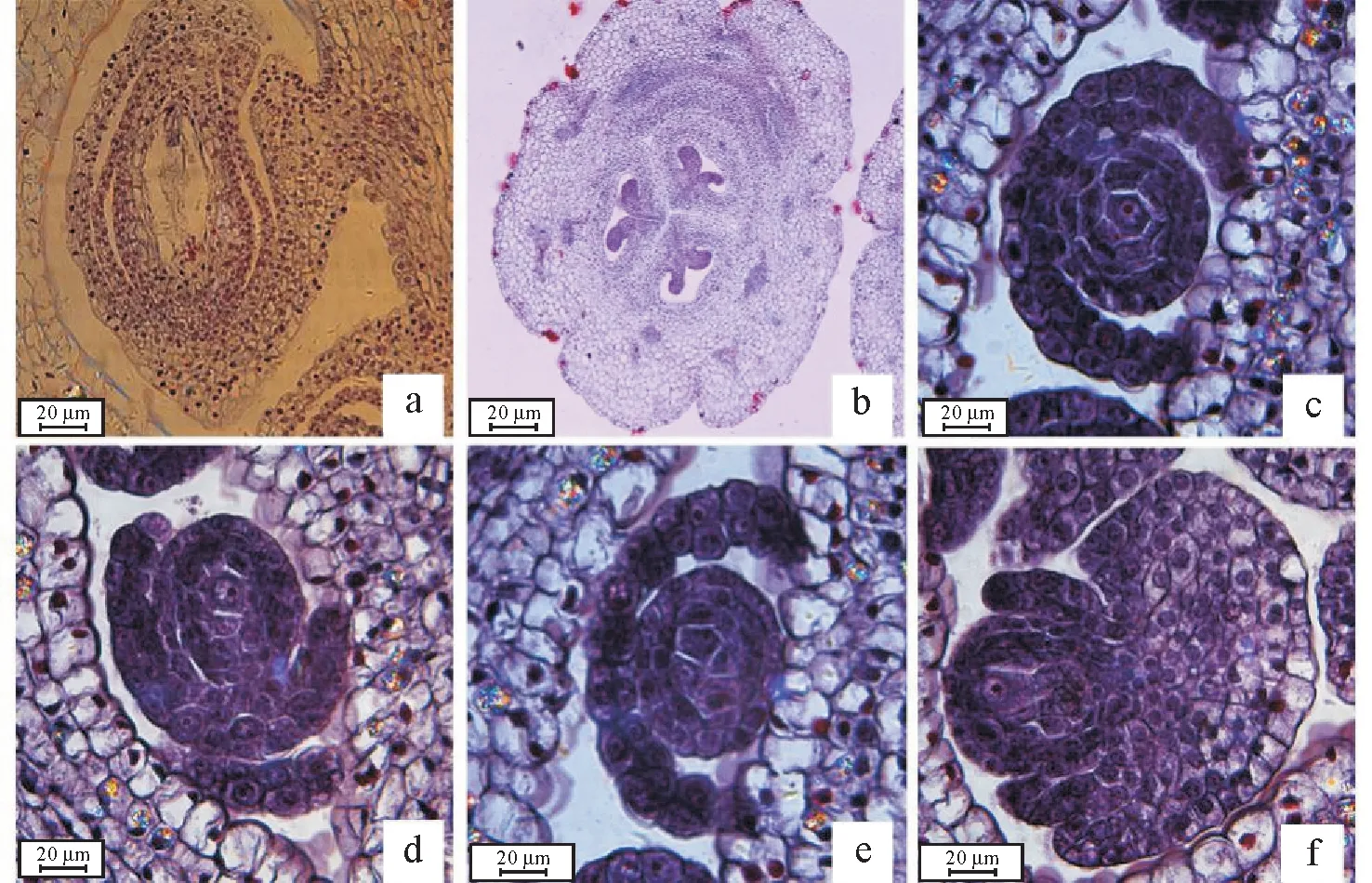
图4 大孢子发生Fig.4 Megasporogenesis
2.2.2 雌配子体形成 大孢子继续发育,核发生一次分裂形成二核胚囊(图5a,5b),两个核分别在胚囊的两侧各进行有丝分裂,形成四核胚囊(图5c),再分裂一次形成八核胚囊;与此同时,细胞质亦发生分裂形成“七胞八核”成熟胚囊,也称雌配子体。在雌配子体形成过程中,部分核未能正常分裂形成八核胚囊,从而不能正常受精座果,或未能形成健康种胚,这可能是圆齿野鸦椿种子发芽率低的主要原因之一。其成熟胚囊为蓼型胚囊。

图5 雌配子体形成Fig.5 Formation of female gametophytes
2.3 圆齿野鸦椿花粉形态观察
由图可知,花药、花粉均为金黄色(图6a)。扫描电镜下可见花药囊以及依附在周围的花粉粒(图6b)。花粉粒平滑(少量干瘪)、赤道面观呈长球形,有3条萌发沟(图6c)。极面观呈3裂圆形(图6d),观察到许多萌发孔,属N3P4C5型花粉。

图6 花粉形态Fig.6 Pollen morphology
3 结论与讨论
3.1 圆齿野鸦椿小孢子和雄配子体形成特点
3.1.1 绒毡层对雄配子体形成的影响 从雄蕊开始形成造孢细胞到形成小孢子母细胞阶段,绒毡层便开始形成营养物质。在花粉成熟前,绒毡层起到提供、运输营养物质的作用,最后解体消失。关于绒毡层与花粉发育异常之间的关系,据张鹏飞等[16]观察小麦发现,绒毡层提前降解导致营养供给不足,小孢子分裂过程中,细胞核未能正常分裂,最终花粉败育。暴会会等[17]开展番茄花粉败育雄性不育研究时发现,绒毡层细胞提前或推迟进行细胞程序性死亡降解为小孢子提供营养物质,都会对雄配子体形成产生影响,DYT1-TDF1-AMS-MS188-MS1 转录因子形成遗传途径控制绒毡层的发育。张江江等[18]进行雄性不育中花粉壁研究时发现,绒毡层降解异常会导致雄性不育的根本原因是因为其会对花粉壁的形成产生影响。EMSI编码存在亮氨酸受体蛋白激酶(LRR-RPK),与绒毡层的发育有关,说明EMSI与花粉壁的发育有关。王馨等[19]在观察檫木雌雄配子体发育时,认为如果花粉在绒毡层运输、提供营养物质之前就已经发育异常,则绒毡层就不用提供营养物质,也就不会出现解体消失。花粉发育异常发生在形成造孢细胞之前,绒毡层也就无需发挥其作用,花粉败育与之无关。本研究中,圆齿野鸦椿雄蕊原基分化开始细胞未出现异常;花粉发育后期,部分绒毡层未解体退化,且花粉出现解体空腔可能与之有关。绒毡层解体异常对小孢子发育产生的影响有待进一步研究。
3.1.2 花粉败育现象 张鹏飞等[20]研究发现花粉败育与很多因素有关;呼吸作用中发挥作用的线粒体出现异常会导致植物细胞死亡;过氧化物酶活性异常与影响蛋白合成之间的关系;多糖、脂类以及蛋白质的异常积累和表达对小孢子的育性产生不良的影响。王馨等[21]研究认为檫木在减数分裂时期,小孢子染色体(染色质)分布不均导致花粉空腔,究其原因应该与相关基因表达有关。本研究中,圆齿野鸦椿仅出现部分花粉粒空腔,整体对受精结实影响不大,空腔花粉形成的具体原因有待进一步研究。
3.2 圆齿野鸦椿大孢子和雌配子体形成特点
3.2.1 胚囊败育现象 张小方[22]研究表明,文冠果大孢子减数分裂未能形成功能大孢子,4 个细胞均退化出现空囊是胚囊败育的主要原因。圆齿野鸦椿在形成功能大孢子时,因细胞核退化出现空囊现象与上述情况类似,可能与大孢子细胞内染色体分裂不均有关。樊丽莉等[23]研究楸树发现,胚珠发育需要大量营养,胚囊败育情况大多在二核向八核分裂的时期。出现败育情况可能是营养分配不均,植物为保全大局而做出的选择。左亚君等[24-25]研究发现,粗柄独尾草在二核胚囊向八核胚囊发育时,亦出现胚囊败育现象。张超越等[26]在西瓜胚胎发育研究中发现,由于养分有限,晚发生的胚珠会选择败育以保证先发生的胚珠正常发育受精。张爱勤等[27]研究发现,豆科植物也经常出现选择性败育,物质资源有限,需确保最大繁殖效率。陈利娜等[28]研究表明,雌蕊败育受珠被和胚囊发育、花柱结构以及整个雌蕊分化过程影响。本研究中,圆齿野鸦椿在花芽分化过程中需要大量营养物质,营养供应不足亦可导致胚囊败育,具体原因需进一步探究。
3.2.2 圆齿野鸦椿落花落果的原因 桂明珠等[29]研究发现,雌雄配子体败育以及植物体内有限营养资源的竞争会导致落花落果。许培磊等[30]研究认为,果树的品种特性、雌雄配子体发育、植物体内源激素变化以及外部天气、营养条件等都会使果树出现落花落果的现象。本研究发现,圆齿野鸦椿出现部分雌雄配子体败育现象;二核胚囊阶段以后,可见卵细胞部分缺失助细胞,受精后未能形成正常种胚;此外,圆齿野鸦椿冬芽多为混合芽,春季叶芽与花芽同时分化,且花量大,花芽分化持续时间长(3 个月左右),需消耗大量营养,故出现营养竞争;这些情况可能导致该树种落花落果。因此,圆齿野鸦椿养护过程中,应及时施肥,尤其应该重视基肥的应用,以确保春季萌芽时养分供应充足,以提高坐果率。
3.3 结论
雄配子体形成过程中,存在部分败育现象,包括花粉粒空腔、绒毡层降解异常。雌配子体形成过程中,存在功能大孢子退化、二核至八核分裂时期出现异常,未能正常分裂形成八核胚囊,从而不能正常受精座果或不能形成健康种胚。该研究解释了圆齿野鸦椿空籽率高、种子发芽率低的原因,为解决花粉不育、受精不亲和,胚胎败育等问题提供理论依据。
