高代转玉米C4-pepc基因水稻株系形态和产量特性及其聚类分析
2021-09-26王净周兴雅张金飞曹悦吴博晗李霞
王净 周兴雅 张金飞 曹悦 吴博晗 李霞
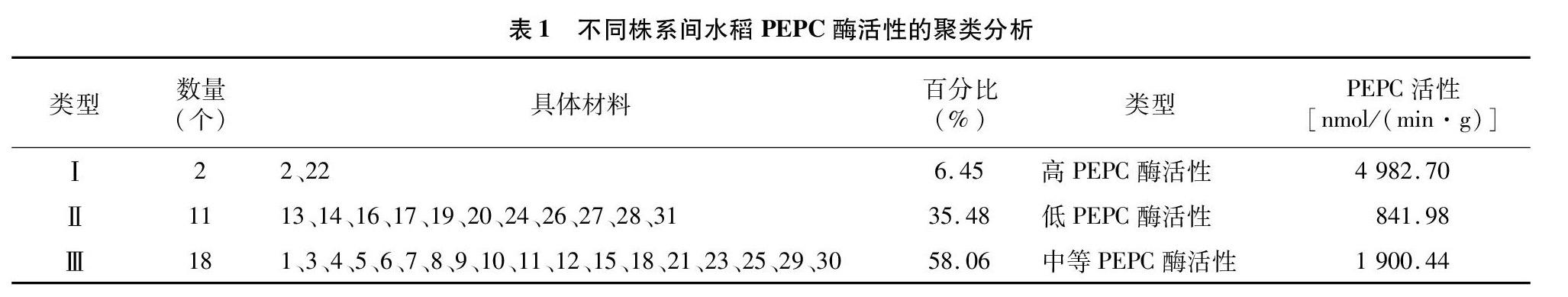


摘要:为获得高光效的转C4型PEPC(磷酸烯醇式丙酮酸羧化酶phosphoenolpyruvate carboxylase,PEPC,EC.1.1.31)基因水稻(简称PC)高代的稳定株系,选取2019—2020年种植的同一株系的PC材料,分析PC材料的形态、光合参数、酶活性以及产量构成因子等的变化特点,并将各参数进行聚类分析。结果表明,PC高代材料的PEPC酶活性、净光合速率以及产量构成因子等指标的确存在年度间和株系间的变化,其中变异系数表现为产量构成因子<净光合速率 关键词:水稻;PEPC酶;净光合速率;产量因子;聚类分析 中图分类号: S511.01 文献标志码: A 文章编号:1002-1302(2021)17-0091-05 收稿日期:2020-12-17 基金项目:国家自然科学基金(编号:31571585);江苏省重点研发计划(编号:BE2019377);国家重点研发计划(编号:2016YFD0300501-03)。 作者简介:王 净(1994—),女,河南周口人,硕士研究生,主要从事水稻作物生理研究。E-mail:1790438668@qq.com。 通信作者:李 霞,博士,研究员,主要从事水稻逆境生理研究。E-mail:jspplx@jaas.ac.cn。 水稻(Oryza sativa L.)是全球一半以上人口的主食,其超高产量品种和栽培技术的发展都建立在株型改良的基础上[1]。利用数量性状(quantitative trait locus,QTL)分析发现,株高主要受顶部1节间和顶部4节间长度的影响[2]。在保证抗倒性和保持收获指数不变的前提下,株高的增加可提高生物量和最终产量[3]。水稻高光效机制是水稻超高产育种的重要理论基础,其中单个C4型磷酸烯醇式丙酮酸羧化酶(phosphoenolpyruvate carboxylase,PEPC,EC.1.1.31)转基因水稻(简称PC)具高光效特性,受到作物科学工作者的广泛关注[4]。 高表达转C4型PEPC水稻在高光强和高温条件下,外源导入的C4-pepc表达和PEPC酶活性均显著增加,而且功能叶片的光合效率也显著提高[5],单株产量比未转基因的野生型原种Kitaake(简称WT)也显著提高[6],并将水稻开花后5~10 d剑叶的PEPC酶活性作为水稻高光效育种筛选的可靠生理指标[4]。分子机制研究发现,PEPC酶活性增强和C4-pepc表达上调与NO和Ca2+参与蛋白激酶和转录因子的介导密切相关[7]。在干旱胁迫下,PC水稻中C4-pepc的调节与其糖信号-蔗糖非发酵蛋白激酶(sucrose nonfermenting-1-related protein kinase,SnRKs)家族的调节有关,如亚家族 1-SnRK1 相关基因OsK1a、OsK24、OsK35的表达和SnRK2均高于WT[8]。而外源葡萄糖施入试验表明,SnRK3s家族基因可与钙调B类磷酸酶(calcineurin-B-like,CBL)作用诱导NO 参与PC叶片气孔调节,从而有利于PC 耐旱[9]。因此,PC作为水稻干旱耐性机制研究材料而被关注。但随着这10余年的研究以及多代繁殖,观察到高代转C4-pepc水稻株系的酶活性表达不稳定,且呈下降趋势,导致相应的转基因材料耐旱优势也受限。因此,如何使外源导入的有益基因在水稻内稳定表达,是有益基因能否为水稻育種工作者利用的前提。前期的研究已观察到不同的测定条件和不同的生育期是影响高代转C4-pepc水稻株系酶活性发挥的重要因子[4-5],但是高代PC 株系在相同条件下仍然存在酶活性的差异,这在一定程度上影响了该材料耐性生理机制的深入研究和材料在育种中的应用。因此,本研究选取不同年份同一株系的高代PC材料,研究年度间高代PC材料形态、光合参数、酶活性以及产量构成因子的变化特点,并将各参数进行聚类分析,以确定稳定的筛选指标,用于高代高光效水稻材料的利用。期望该研究可为筛选高效稳定的高光效水稻材料提供参考,也可为今后解析其高光效差异的内在机制提供新视角。 1 材料与方法 1.1 试验材料 本研究选取的PC高代材料最初由美国华盛顿州立大学古森本教授赠送,该材料是通过农杆菌介导获得高表达转玉米C4-pepc全基因的第3代水稻株系,获得的水稻种子每年都在江苏省农业科学院转基因作物基地种植。在本研究中,选取2018—2019年种植的高代材料31个株系,每个株系都是上一年收集的单穗种子于次年种植获得的植株。水稻种子分别于2019年、2020年的5月1日播种,在水泥池种植,每穴播1苗,每个单穗种植的材料作为1个株系,并编号、挂牌,常规水肥和病虫草害管理,于7月底在植株开花后7~14 d,测定剑叶的光合参数,同步取样,液氮保存,用于测定叶片的酶活性,成熟后单株收获。 1.2 光合参数测定 参照王超等的方法[5],使用美国LI-COR公司生产的LI-6400便携式光合测定仪,开放式系统红蓝光源测定。晴天的08:30—11:30,室外温度25~30 ℃,相对湿度70%~80%,测定水稻株系剑叶的净光合速率(net photosynthetic rate,Pn),室外进行,每个处理5~6次重复。 1.3 形态指标的测定 在开花后50 d收获植株,每个株系在收获时选取5株测定其形态指标,包括株高、有效分蘖数和穗长等。 1.4 叶片磷酸烯醇式丙酮酸羧化酶(PEPC)活性的测定 参照 Giglioli等的方法[10]测定PEPC的活性。简单来说将0.1 g水稻剑叶置于提前冷冻的研钵中研磨,加入1 mL粗酶提取缓冲液,在4 ℃下 13 000 g 离心10 min,收集上清液(即粗酶液)。在酶活性测定的反应体系(1 mL)中依次加入725 μL混合液,25 μL 5 mmol/L 磷酸烯醇式丙酮酸(phosphoenolpyruate,PEP),150 μL粗酶液摇匀后,用紫外分光光度计(UV-1200型,上海美谱达仪器有限公司),记录340 nm处吸光度的变化,计算酶活性。 1.5 产量构成因子测定 在开花后40 d,每个株系选3株长势一致的供试材料单株收获,自然晾晒至籽粒含水量小于14%时,在室内进行考种,并测定其产量构成因子。 1.6 数据处理 使用SPSS 22.0软件对数据进行统计以及聚类分析,采用Microsoft excel 2016软件对数据进行描述和作图。 2 结果与分析 2.1 高代PC株系剑叶PEPC酶活性的变化及其聚类分析 水稻生育后期剑叶的PEPC酶活性为水稻高光效特性的重要筛选指标[4]。因此,本研究选取了高代PC 株系共31株,连续2年选取水稻株系的剑叶测定其PEPC酶活性,所有株系PEPC 酶活性为(1 723.72±1 139.14) nmol/(min·g),变异系数为0.66。并使用WARD 法对PEPC酶活性数据进行聚类分析 (度量标准为Euclidean 距离),以阈值 20 作为聚类标准,可将供试材料的类型量化细分为3类(表1)。其中类型Ⅰ为高PEPC酶活性类型,PEPC酶活性为4 982.70 nmol/(min·g),包括第2、22株系,占供试材料的 6.45%;类型Ⅱ为低PEPC酶活性类型,PEPC酶活性为 841.98 nmol/(min·g),包括第13、14、16、17、19、20、24、26、27、28、31株系,占供试材料的35.48%;类型Ⅲ 为PEPC酶活性位于中间的材料类型,PEPC酶活性为1 900.44 nmol/(min·g),包括第1、3、4、5、6、7、8、9、10、11、12、15、18、21、23、25、29、30株系,占供试材料的58.06%。可见,高代PC的PEPC酶活性的确存在一定的变异。 2.2 高代PC株系剑叶净光合速率的变化及其聚类分析 连续2年同步测定对应株系抽穗期,开花后 14 d 剑叶的净光合速率(photosyntheticrate,Pn),2年所有株系的平均值和标准差为(17.11±4.14) μmol/(m2·s),变异系数为0.24,将该参数经过WARD法聚类分析(度量标准为Euclidean距离),以阈值8作为聚类标准,也可将供试材料的类型量化细分为3类。其中类型Ⅰ为高净光合速率类型,包括第1、2、3、7、22、23株系,占供试材料的19.35%,净光合速率平均值为 21.53 μmol/(m2·s);类型Ⅱ为低净光合速率类型,包括第15、20、21、29、30株系,占供试材料的16.13%,净光合速率的平均值为 14.65 μmol/(m2·s);类型Ⅲ为净光合速率位于中间的材料类型,包括第4、5、6、8、9、10、11、12、13、14、16、17、18、19、24、25、26、27、28、31株系,占供试材料的64.52%,净光合速率为 16.40 μmol/(m2·s),和酶活性的表现一样也呈现正态分布(表2)。 2.3 高代PC株系产量构成因子的变化及其聚类分析 连续2年在同期同步记录了不同株系产量构成因子的变化,其中2年的平均值分别为株高为(75.38±3.64) cm,变异系数为0.05;总分蘖数为(9.85±1.84)个,变异系数为0.19;有效分蘖数为(7.64±1.56)个,变异系数为0.20;千粒质量为(21.88±0.58)g,变异系数为0.03;穗长为(10.49±0.65) cm,变异系数为0.06;穗粒数为(28.32±4.05)个,变异系数为0.14;结实率为(72.21±6.85)%,变异系数为0.09;单株产量为(3.56±1.19) g,变异系数为0.33。可见,供试材料各形态指标变异小于其单株产量,单株产量的变化主要与有效穗数和每穗粒数的变化密切相关。进一步将这些数据经过WARD法聚类分析(度量标准为Euclidean距离),以阈值12作为聚类標准,也可将供试材料的类型量化细分为3类。其中类型Ⅰ为高产类型,包括第4、5、7、13、16、17、21、22、30株系,占供试材料的29.03%;类型Ⅱ为低产类型,包括第2、6、12、20、23、25、26、27、28、31株系,占供试材料的32.26%;类型Ⅲ为产量位于中间的材料类型,包括第1、3、8、9、10、11、14、15、18、19、24、29株系,占供试材料的38.71%(表3)。 2.4 PEPC酶活性、净光合速率和产量指标的聚类分析 进一步将本研究中PC高代材料测定的3类指标数据也经过WARD法聚类分析(度量标准为Euclidean距离),以阈值15作为聚类标准,可将供试材料的类型量化细分为3类(表4)。其中类型Ⅰ为高PEPC酶活性,高净光合速率和中产的材料类型,包括第1、2、3、7、22、23株系,占供试材料的19.35%;类型Ⅱ为低PEPC酶活性,低净光合速率和高产的材料类型,包括第13、14、16、17、19、20、24、26、27、28、31株系,占供试材料的35.48%;类型Ⅲ为中等光合能力的材料类型,包括第4、5、6、8、9、10、11、12、15、18、21、25、29、30株系,占供试材料的45.16%。从图1可以看出,类型Ⅰ实际上还可以细分为2个小的亚型,Ⅰ-1包括1、3、7、23共4个株系,占供试材料的12.90%,这类型的PEPC活性[平均值2 747.19 nmol/(min· g)]和净光合速率[平均值为21.49 μmol/(m2·s)]仍高于其他2个大类型(Ⅱ和Ⅲ),而且单株产量最高(平均值为4.18 g/株);而Ⅰ-2,只包括2个株系(2和22),占比6.45%,这类型的PEPC酶活性最高[平均值 4 982.70 nmol/(min· g)],净光合速率与Ⅰ-2的类似,也较高[平均值为 21.60 μmol/(m2·s)],但是产量则最低(平均值为2.87 g/株)。进一步从各类参数的相关性分析可知,PC株系的Pn与PEPC酶活性的相关系数为0.808,呈极显著相关(P<0.01),PEPC酶活性的发挥与光合能力密切相关,但是与单株产量相关性不显著。 3 讨论与结论 C4植物与C3植物的一个重要区别是C4植物的CO2补偿点很低,所以C4植物在CO2含量低的情况下存活率更高,而且C4植物在高光强、高温和干旱等条件下,比C3植物具有较高的CO2固定效率、水分利用率和籽粒產量[11],其中C4植物光合作用的关键酶PEPC起关键作用。科学家已经通过基因工程,将该基因引入C3植物如水稻中,该基因不仅在水稻中获得高表达,而且表现高光效,并最终表现出籽粒产量提高[12-13]。与原种Kitaake相比,高表达转玉米pepc基因水稻(PC)不仅表现出较强的光合能力和较高的籽粒产量,而且还在强光、高温、干旱和低氮下均具有较高的净光合速率和籽粒产量,并与高PEPC酶活性的诱导密切相关[8,14-18],可以作为研究植物耐逆机制的有益资源。 磷酸烯醇式丙酮酸羧化酶在 C4植物中起浓缩 CO2以提高光合效率的作用。王强等研究表明,两优培九超高产的重要原因,与其较高的光能和CO2利用效率、较强的抗光抑制能力以及较高的 C4途径酶PEPC的表达水平相关[19]。研究表明,PEPC酶在不同生育期、不同部位以及不同光强条件下的表达特征曲线与光合速率具有密切关系,特别在产量形成的关键时期,外源PEPC基因高水平表达与净光合速率的高值出现期表现一致,而且相对稳定,此时测定的剑叶PEPC活性可作为水稻高光合速率的可靠分子育种指标,这为水稻高光效特性鉴定和筛选提供了依据,并能为作物高光效机制的研究提供有用的材料[4]。本研究利用相关的筛选指标,检测了年度间高代的31株系PC水稻的高光效特性和产量表现。结果表明,PC的高代材料在PEPC酶活性的发挥、净光合速率以及产量构成因子等指标上的确存在年度间和株系间的变化,其中变异系数产量构成因子<净光合速率 磷酸烯醇丙酮酸羧化酶包括植物型PEPC(plant-type phosphoenolpyruvate carboxylase,PTPC)和细菌型PEPC(bacterial-type phosphoenolpyruvate carboxylase,BTPC)等多基因家族。BTPC和PTPC相互作用,形成2类异性寡聚多肽复合物[20],在蓖麻(Ricinus communis)种子发育中发挥重要作用。因此,它的功能很多样,而且PEPC酶和基因的调节也很复杂。前期研究表明,与WT相比,PC材料在高光强[14]、高温[15]、干旱[7]以及低氮条件[17]下均表现出较好的耐性,而且这些优异的特性与PEPC酶活性的发挥以及C4-pepc基因的调节密切相关。信号分子H2O2[6]、NO[21]、Ca2+[7]、磷脂酸[22]、 ATP[23] 以及糖分子[24-25]均参与其基因的表达。本研究还观察到PC水稻在响应干旱逆境时C4-pepc有去甲基化现象发生,有利于C4-pepc的高表达,也伴随着PEPC的磷酸化以及相关基因的表达,显示了表观遗传机制可能也参与该材料干旱的响应机制[8]。因此,深入研究PC不同株系PEPC酶活性的调节机制,是获得稳定高光效材料的重要理论基础,将有助于揭示其基因表达差异的内在原因。 参考文献: [1]Tian Z X,Wang J W,Li J Y,et al. Designing future crops:challenges and strategies for sustainable agriculture[J]. The Plant Journal:for Cell and Molecular Biology,2021,105(5):1165-1178. [2]姜树坤,黄 成,徐正进,等. 粳稻株高QTL与赤霉素和油菜素内酯合成及信号转导基因相关分析[J]. 中国农业科学,2010,43(14):2829-2838. [3]Zhu X G,Long S P,Ort D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology,2010,61(1):235-261. [4]李 霞,王 超,陈 晏,等. PEPC酶活性作为水稻高光效育种筛选指标的研究[J]. 江苏农业学报,2008,24(5):559-564. [5]王 超,李 霞,蔡庆生. 不同测定环境条件下转PEPC基因水稻及杂交后代光合特性的比较[J]. 江苏农业学报,2008,24(3):232-236. [6]Ren C G,Li X,Liu X L,et al. Hydrogen peroxide regulated photosynthesis in C4-pepc transgenic rice[J]. Plant Physiology and Biochemistry,2014,74:218-229. [7]Qian B Y,Li X,Liu X L,et al. Improved oxidative tolerance in suspension-cultured cells of C4-pepc transgenic rice by H2O2 and Ca2+under PEG-6000[J]. Journal of Integrative Plant Biology,2015,57(6):534-549. [8]Liu X L,Li X,Zhang C,et al. Phosphoenolpyruvate carboxylase regulation in C4-pepc expressing transgenic rice during early responses to drought stress[J]. Physiologia Plantarum,2017,159(2):178-200. [9]張金飞,李 霞,何亚飞,等. 外源葡萄糖增强高表达转玉米C4型pepc水稻耐旱性的生理机制[J]. 作物学报,2018,44(1):82-94. [10]Giglioli-Guivarch N,Pierre J N,Brown S,et al. The Light-Dependent transduction pathway controlling the regulatory phosphorylation of C4 phosphoenolpyruvate carboxylase in protoplasts from Digitaria sanguinalis[J]. The Plant Cell,1996,8(4):573-586. [11]Zhu X G,Long S P,Ort D R. Improving photosynthetic efficiency for greater yield[J]. Annual Review of Plant Biology,2010,61(1):235-261. [12]Ku M S,Agarie S,Nomura M,et al. High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants[J]. Nature Biotechnology,1999,17(1):76-80. [13]Taniguchi Y,Ohkawa H,Masumoto C,et al. Overproduction of C4 photosynthetic enzymes in transgenic rice plants:an approach to introduce the C4-like photosynthetic pathway into rice[J]. Journal of Experimental Botany,2008,59(7):1799-1809. [14]Jiao D M,Ji B H,Li X. Characteristics of chlorophyll fluorescence and membrane-lipid peroxidation during senescence of flag leaf in different cultivars of rice[J]. Photosynthetica,2003,41(1):33-41. [15]李 霞,焦德茂,戴传超. 转PEPC基因水稻对光氧化逆境的响应[J]. 作物学报,2005,31(4):408-413. [16]唐玉婷,李 霞,陆 巍,等. 高表达转 C4型PEPC基因水稻在低氮下诱导碳氮酶稳定光合作用[J]. 华北农学报,2015,30(4):95-100. [17]Tang Y T,Li X,Lu W,et al. Enhanced photorespiration in transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase gene contributes to alleviating low Nitrogen stress[J]. Plant Physiology and Biochemistry,2018,130:577-588. [18]严 婷,李佳馨,李 霞,等. 转C4型PEPC基因水稻非生物胁迫耐受性研究进展[J]. 淮阴工学院学报,2019,28(5):62-68. [19]王 强,卢从明,张其德,等. 超高产杂交稻两优培九的光合作用、光抑制和C4途径酶特性[J]. 中国科学(C辑),2002,32(6):481-487. [20]OLeary B,Park J,Plaxton W C. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase):recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs[J]. The Biochemical Journal,2011,436(1):15-34. [21]Chen P B,Li X,Huo K,et al. Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors[J]. Journal of Plant Physiology,2014,171(6):458-466. [22]Li X,Wang C,Ren C G. Effects of butanol neomycin and Calcium on the photosynthetic characteristics of pepc transgenic rice[J]. African Journal of Biotechnology,2011,76(10):17466-17476. [23]Huo K,Li X,He Y F,et al. Exogenous ATP enhance signal response of suspension cells of transgenic rice (Oryza sativa L.) expressing maize C4-pepc encoded phosphoenolpyruvate carboxylase under PEG treatment[J]. Plant Growth Regulation,2017,82(1):55-67. [24]Zhang C,Li X,He Y,et al. Physiological investigation of C4 phosphoenolpyruvate carboxylase introduced rice line shows that sucrose metabolism is involved in the improved drought tolerance[J]. Plant Physiology and Biochemistry,2017,115:328-342. [25]He Y F,Xie Y F,Li X,et al. Drought tolerance of transgenic rice overexpressing maize C4-pepc gene related to increased anthocyanin synthesis regulated by sucrose and calcium[J]. Biologia Plantarum,2020,64:136-149.
