我国鱼类胚胎发育研究进展
2021-09-26陈军平沈方方武慧慧张佳鑫于若梦付永杰谢国强赵道全
陈军平 沈方方 武慧慧 张佳鑫 于若梦 付永杰 谢国强 赵道全

摘要:鱼类胚胎发育一直都是水产育种工作中最为关键的一个环节,对种群延续及渔业资源可持续发展也至关重要。而鱼类胚胎在发育过程中极易受外界环境的影响,进而对胚胎受精率和孵化率有较大影响,因此需要对不同鱼类胚胎发育的生物学特征进行深入研究。本研究通过对我国近些年来鱼类胚胎发育的研究现状进行总结,分析探讨了孵化温度、盐度及其他因素对我国鱼类胚胎发育的影响,最后也探讨了我国杂交鱼类胚胎发育的研究近况,以期为我国更多鱼类繁育研究提供一定参考。
关键词:鱼类;胚胎发育;温度;盐度;杂交;研究进展
中图分类号: S917 文献标志码: A
文章编号:1002-1302(2021)17-0045-08
收稿日期:2021-01-19
作者简介:陈军平(1993—),男,河南周口人,硕士研究生,实习研究员,主要从事鱼类遗传育种研究。E-mail:chenjunpinghn@163.com。
通信作者:赵道全,教授级高级工程师,主要从事冷水鱼、工厂化养鱼和种质资源保护等研究。E-mail:Zhaodaoquan@163.com。
鱼类胚胎发育不仅是动物遗传和发育生物学中重要的研究课题,也是水产育种工作中最为关键的一个环节。胚胎发育是从一个受精卵不断发育成比较成熟且形态正常、健康个体的过程。一直以来,众多学者使用不同方法对鱼类胚胎发育的各个生物学特征进行深入研究,大大提高了我国相关鱼类的孵化率和成活率,为我国鱼类发育学、鱼类种质资源保护打下坚实的基础。本研究根据我国相关鱼类胚胎发育的研究现状,分析探讨了孵化温度、盐度及其他因素对我国鱼类胚胎发育的影响,最后也探讨了我国杂交鱼类胚胎发育的研究近况,以期为我国更多鱼类(稀有鱼类)繁育,种质资源开发、利用和保护提供理论依据,促进我国渔业资源又好又快发展。
1 胚胎发育
与其他动物一样,鱼类胚胎在整个发育过程中变化最为明显,不论其所处环境还是所属种类,均容易受外界坏境不同程度的影响,对受精率、畸形率和成活率均有较大影响,所以这一阶段对于鱼的个体来说既是重要的又是脆弱的。即使基因对鱼类胚胎发育有着决定性作用,但外界因素如温度、盐碱度、溶解氧和pH值等也会对胚胎的整体发育过程产生巨大的影响[1]。由于胚胎容易受到外界各因素的影响,因此在各生长阶段中死亡率也是最高的,这就迫切需要根据不同种属胚胎发育的特点进行更深入研究,以期更好地保证鱼类胚胎孵化率和成活率,这对于未来更高效的工厂化、集约化及渔业种质资源保护和渔业可持续发展具有重要意义。
硬骨鱼类的胚胎发育主要经历5个时期,分别为卵裂期、囊胚期、原肠胚期、神经胚期和器官形成期[2]。目前,国内学者已对大量鱼类胚胎发育情况进行了比较深入的观察,发现了温度等环境因素对其胚胎发育速率均有不同程度影响,如虹鳟(Oncorhynchus mykiss)[3]、黄姑鱼(Nibea albiflora)[4]、拟鲿(Pseudobagrus ussuriensis)[5]、斑点叉尾(Ictalurus punctatus)[6]、黄斑篮子鱼(Siganus oramin)[7]、黄尾鲴(Xenocypris davidi Bleer)[8]等。
2 环境因素对鱼类胚胎发育的影响
2.1 温度对胚胎发育的影响
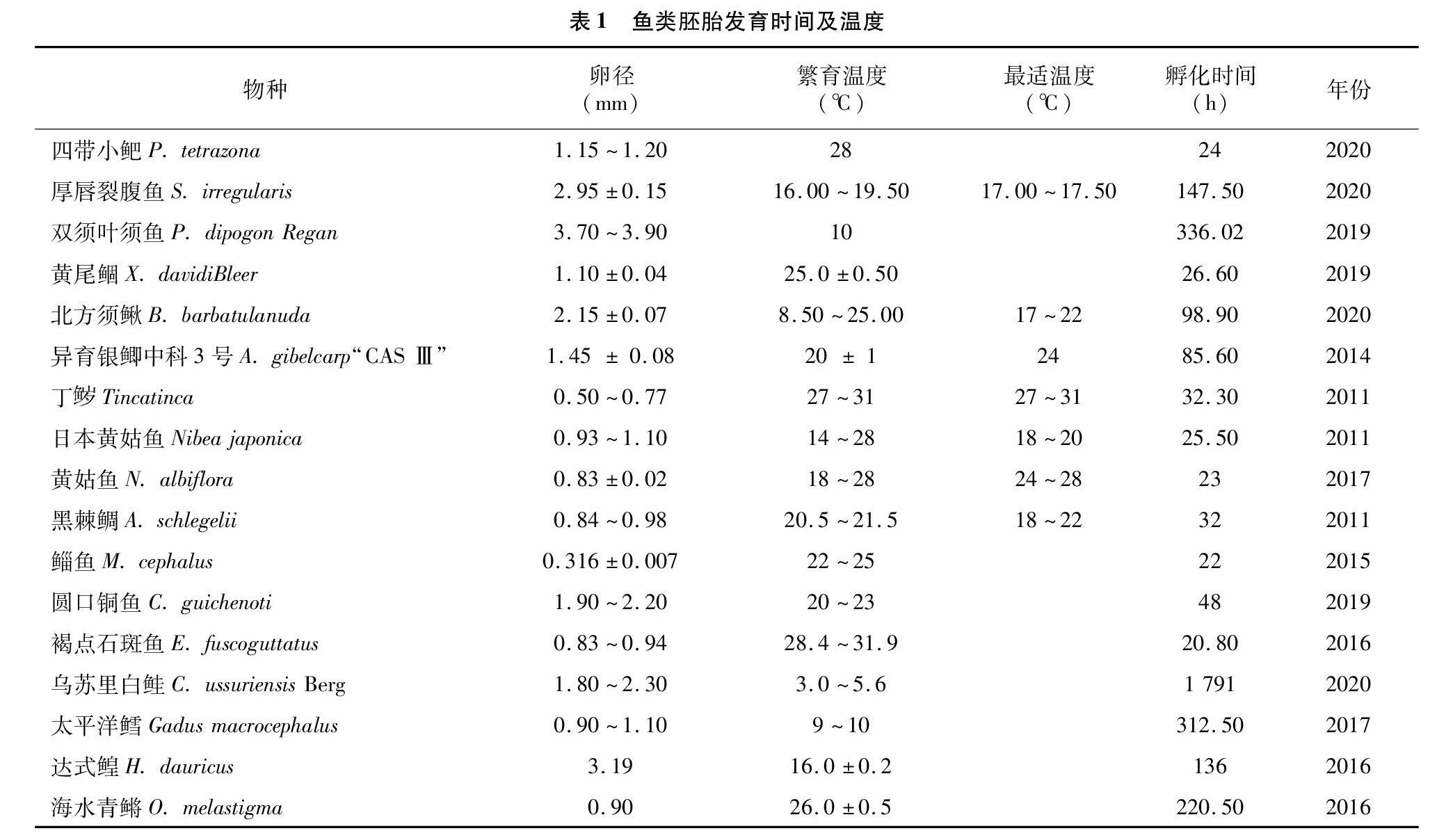
温度对亲本产卵、胚胎发育、后期幼鱼的生长过程及鱼类的摄食和新陈代谢等生理过程均有重要影响。其中,胚胎发育过程对水温的高低最为敏感[9]。在鱼类胚胎发育过程中,温度过高或过低均会导致胚胎停止发育或死亡,造成孵化率降低等,所以研究最适胚胎发育温度尤为重要。近几年,育种工作者对越来越多的鱼类进行了早期胚胎发育的研究,发现水温对胚胎发育的各个阶段及孵化率有较大的影响,如圆口铜鱼(Coreius guichenoti)[10]、鲻鱼(Mugil cephalus)[11]、胭脂鱼[Myxocyprinus asiaticus (Bleeker)][12]、半滑舌鳎(Cynoglossus semilaevis)[13]、达式鳇(Huso dauricus)[14]、褐点石斑鱼(Epinephelus fuscoguttatus)[15]、黄颡鱼(Pelteobagrus fulvidraco)[16]等。其中,根據孵化温度变化对胚胎发育的影响,已确定了异育银鲫“中科 3 号”(Allogynogenetic gibelcarp “CAS Ⅲ”)[17]、北方须鳅(Barbatulabar batulanuda)[18]、丁(Tinca tinca)[19]等鱼类胚胎发育的最适孵化温度,大大提高了相关鱼类的繁育效率(表1)。郭永灿发现草鱼(Ctenopharyngodon idella)和鲢(Hypophthalmichthys molitrix)的胚胎在16~33 ℃才能正常存活,在短时间内温度上升或下降3~5 ℃时,草鱼卵和鲢卵在囊胚期和原肠期会产生一些不良反应,尤以原肠期最为明显,造成孵化率大幅度下降甚至鱼卵死亡[20]。
对于同是鲤科鱼类的尖裸鲤(Oxygymnocypris stewartii)[21]和四川华鳊(Sinibrama taeniatus)[22]来说,在不同温度条件下,胚胎发育时间与温度呈显著性负相关函数关系,尖裸鲤的胚胎在水温8 ℃时受精率最高,可达100%,而在17 ℃时最先发育到原肠期且孵化时间最短。四川华鳊胚胎在16 ℃时发育至原肠期时出现全部死亡现象,与大多数鱼类一样,其孵化时间随着温度上升而缩短,但其受精率和孵化率均随着温度的上升出现了先上升后下降的现象,在25 ℃时孵化率最高为96.13%,在 31 ℃ 时孵化率为79.11%,畸形率也高达10.49%,该结果也与相关研究结果[23]一致。黄贤克等在观察黄姑鱼胚胎发育时发现,当水温在18~28 ℃之间时对黄姑鱼孵化率和畸形率影响较小,随着温度的升高,缩短了孵化时间,当孵化水温为26 ℃时,黄姑鱼胚胎的器官形成期明显缩短,说明这一时期对温度非常敏感,适宜胚胎器官的形成[24],其最适水温为 20~24 ℃与黄永春等对状黄姑鱼(Nibea miichthioides)胚胎发育的研究结果[25]一致。大多数鱼类在胚胎发育过程中都会不同程度受到外界环境变化的影响,也就是“敏感期”,但史秀兰等发现乌苏里白鲑(Coregonus ussuriensis Berg)在胚胎发育中并没有出现“敏感期”,而其他鲑科鱼类在胚胎发育中均出现敏感期,在硬骨鱼类中,先天免疫系统易受到温度的影响,这也有可能是由于孵化温度不同造成的[26]。
一直以来,斑马鱼都被作为模式生物来开展实验研究,但斑马鱼在出膜时会形成色素,这样并不利于观察早期器官的发育情况。而崔文耀等研究发现,四带小鲃(Puntius tetrazona)胚胎在水温28 ℃时经过24 h就能孵化出膜,其出膜时没有相关色素的形成,这样更利于观察鱼类早期器官形成发育情况,其全年可繁殖、产卵较多,胚胎发育速度也比斑马鱼更快[27],是一种有发展前景的模式生物[28]。王友红等观察海水青鳉(Oryzias melastigma)胚胎发育时发现,其历经312 h后陆续孵化出膜[29],在水温相似的情况下,该研究孵化时间比陈漪等的研究结果[30]慢了近92 h,这可能是由于水质不同等相关因素的原因,影响了其胚胎发育。作为现已报道裂腹鱼类卵径最大的双须叶须鱼(Ptychobarbus dipogon Regan),其卵径为3.7~3.9 mm,孵化用时为336.02 h,远远慢于厚唇列腹鱼胚胎发育时间,与其他裂腹鱼亚科鱼类胚胎发育温度相比偏低,所以其孵化时间较长不仅与温度有关系,还与其卵径较大有很大的关系[31]。在相同的条件下,受精卵越大,其发育速率越慢。官曙光等发现,黑棘鲷(Acanthopagrus schlegelii)的胚胎发育最适温度为 18~22 ℃,当孵化温度高于25 ℃时,在囊胚期和原肠晚期会出现大量死亡现象,这也可能是由于高温影响细胞分裂造成的[32]。相关文献表明,鱼类胚胎发育阶段把水温设定为恒定温度并不是最适温度,而根据胚胎不同发育阶段设定不同的温度,将会更有利于提高胚胎的发育速率,进而提高鱼卵的孵化率和成活率[33-34]。关于不同水温对淇河鲫(Carassius auratus)早期胚胎发育的影响中,吴利敏等发现当孵化早期水温为24 ℃时可提高受精率,后期使水温控制到27 ℃时,可大大提高孵化率[35],这也表明在鱼类胚胎不同发育时期设定不同的温度,将会更有利于胚胎发育。上述研究结果表明,在一定温度范围内,随着温度的升高,鱼类胚胎孵化用时越短;鱼卵卵径越大,孵化用时越长,卵径越小,孵化用时越短;大多数鱼类胚胎发育至原肠期时对温度变化最为敏感。
2.2 盐度对胚胎发育的影响
与温度一样,盐度在鱼类胚胎发育过程中发挥着重要作用,尤其是对海水鱼类影响较大,过高或过低的盐度都会造成受精卵卵膜渗透压的不同,进而使细胞分裂不正常,最终导致胚胎死亡或出膜后出现畸形的现象[36]。周天舒等研究了不同鹽度对大鳍弹鱼(Periophthalmus magnuspinnatus)胚胎的耐受性发现,当盐度高达30‰和35‰时,胚胎发育至囊胚期和原肠晚期出现了全部死亡的现象;在盐度为20‰和25‰时,胚胎均在鳔雏形期死亡;在盐度5‰~15‰的范围内均能正常生长发育,当盐度为10‰时,其孵化率最高,而盐度为5‰时,孵化用时最短。黄贤克等研究发现,黄姑鱼胚胎在盐度为 20‰~24‰之间时,受精卵全部沉底且孵化率较低,由于受精卵全部沉入水底并与死卵接触,部分受精卵得不到充足的氧气进而出现发霉的现象,最终导致孵化率降低[24]。当盐度分别为26‰~32‰时,大部分受精卵悬浮于水中,一小部分受精卵漂浮在水面,且孵化率显著提高,尤其是当盐度为32‰时,孵化率最高达到99.67%,这也是由于受精卵悬在水中或漂浮于水面增加了受精卵在水中的接触面积,能够得到充足的氧气且能较好地排除氨氮,从而大大提高了孵化率。当盐度超过36‰或低于24‰时,孵化出的仔鱼会出现较严重的畸形现象[24],该试验结果与柴学军等对日本黄姑鱼(Nibea japonica)的研究结果[4]相类似。与黄姑鱼相似,条石鲷(Oplegnathus fasciatus)在胚胎发育过程中,当盐度低于26.4‰时,鱼卵只有一小部分漂浮在水的中层,大多数鱼卵沉入底部,高于26.4‰时,鱼卵几乎全部漂浮在水的上层;当盐度低于20.7‰时,受精卵无法正常孵化,当盐度在20.7‰~37.9‰之间时,条石鲷胚胎的孵化率呈不断上升趋势且畸形率不断下降,当盐度高于41.2‰时,其孵化率和仔鱼的成活率均呈下降趋势[37]。蔡文超等还发现,条石鲷胚胎在卵裂时期至原肠期对盐度变化最为敏感,在该时期内均造成大量鱼卵死亡,由于过高或过低的盐度变化导致卵膜内外渗透压不同,从而引起畸形或死亡[38]。宋振鑫等研究观察不同盐度条件对云纹石斑鱼(Epinephelus moara)胚胎发育影响时,发现其胚胎发育在盐度为10‰~45‰时均能正常孵化,其孵化时间基本一致,最适宜盐度在30‰~35‰之间,在该盐度范围内孵化率最高且畸形率最低[39]。该研究结果与七带石斑鱼(Epinephelus septemfasciatus)胚胎发育的最适盐度一致,但是七带石斑鱼胚胎在发育过程中随着盐度的增加,其孵化时间有不断减少的趋势[40]。相关报道称,海水鱼类在胚胎发育过程中受到外界的高强度渗透压时,会在一定程度上促进胚胎发育,进而缩短了孵化时间,但随着盐度的不断增加也会造成受精卵无法正常孵化或孵化后仔鱼出现畸形、抵抗力差等现象[41]。上述研究结果表明,不同鱼类胚胎对盐度的耐受力不同,最适盐度也不相同,适宜的盐度可以使胚胎悬浮于水面,从而得到充足的氧气并提高了孵化率。当盐度过高或过低时,鱼类胚胎均无法孵化出膜,大多数鱼类胚胎发育至囊胚期或原肠期已死亡或停止发育,这也可能是因为胚胎为了调节适应较高的渗透压而消耗了过多能量,从而导致胚胎无法正常发育。
2.3 其他因素对胚胎发育的影响
近些年,由于自然资源和水资源不断遭到破坏,水体中的污染物经过大量积累对鱼类正常发育产生了重大的不良影响。尤其鱼类在早期胚胎发育时期对外界影响因子最为敏感,极易影响鱼类胚胎受精率、发育速率、孵化率,最终导致鱼类中毒、畸形、死亡等现象,而水中的重金属元素严重影响了鱼类早期胚胎的正常发育。史晋绒等研究观察不同浓度的Cu2+对稀有鲫(Gobiocypris rarus)胚胎发育的影响时发现,各个浓度的Cu2+均对稀有鲫有一定毒性影响,出现胚胎心跳减缓、心率不齐、生理功能不正常以及出膜后的仔鱼较小等不良现象。当Cu2+浓度为0.1 mg/L和0.2 mg/L时,其胚胎分别在96 h和60 h全部死亡[42]。然而,一定浓度的CuInS2/ZnS QDs会诱导Hsp70、Cypla和Wnt8a基因表达量上调,使Wstn基因表达发生改变,导致稀有鲫胚胎发育时间延长、孵化率降低和脊柱弯曲等现象,但致死率并不显著[43]。有报道称重金属最早在斑马鱼胚胎发育至卵裂期就出现了畸形现象,斑马鱼受精卵经Zn暴露后,其胚盘就有异常细胞突起等不良现象[44]。黄伟还发现,褐牙鲆(Paralichthys olivaceus)受精卵经重金属Hg和Zn暴露后,其孵化率大大降低[45]。
除此之外,其他影响因素也会对鱼类胚胎发育造成不良影响,当易溶于水的十二烷基苯磺酸钠(SDBS)浓度≥2.5 mg /L时,大鳞副泥鳅(Paramisgurnus dabryanus)胚胎孵化时间显著延长,孵化率明显下降,畸形率较高,这与SDBS抑制了机体抗氧化作用并影响了DNA表达有密切关系,死亡率与SDBS浓度成正比。当SDBS≥0.1 mg/L时,会影响到大鳞副泥鳅精巢发育[46]。在池塘养殖中,残留饵料和鱼类排泄物等原因均会不同程度地导致氨氮含量增高。当氨、氮在一定浓度范围内,胚胎可以正常发育,甚至可以加快胚胎发育速率,但高浓度的氨、氮对鱼类胚胎发育有明显的毒性作用,即使有部分胚胎可以正常出膜,但孵化率会大幅度下降,仔鱼也会出现畸形或无法正常存活[47]。当氨氮浓度为80 mg/L时,台湾泥鳅孵化率会显著下降,即使仔鱼出膜后也会很快死亡,该研究结果与氨氮对鲢、鳙等鱼类影响的结果[48]一致,而不同浓度氨氮对台湾泥鳅胚胎孵化时间及胚胎心率并没有显著影响[49]。我国学者在长江宜昌段对稀有鲫进行暴露试验时发现,当氨氮含量超标时会对稀有鲫胚胎产生较大的毒性效应,造成胚胎心率下降、脊椎弯曲、死亡率高等不良现象[50]。
在养殖过程中,一些兽药、饲料添加劑都曾检测出相关抗生素,这些抗生素并不能完全被鱼类所吸收,造成了严重的环境问题,对鱼类也会产生一定的毒性。刘丽丽等研究发现,当磺胺二甲嘧啶(sulfamethazine,SMZ)为低浓度(0.001~10 mg/L))时,斑马鱼胚胎会出现比较明显的畸形现象,对死亡率影响不明显;SMZ为中浓度(10~300 mg/L)时,对斑马鱼胚胎发育有一定的促进作用,死亡率明显下降;当SMZ为高浓度时,会抑制胚胎正常发育导致死亡率上升,研究表明SMZ对斑马鱼的致畸效应高于致死效应[51]。在用不同浓度的盐酸四环素(tetracycline hydrochloride,TCH)处理斑马鱼母本后,发现其子代胚胎随着母本TCH浓度的升高,孵化率显著降低,畸形率和死亡率均有所增加[52]。当斑马鱼胚胎受精96 h后,经2%乙醇处理后,其孵化率仅为15%,乙醇浓度限制在1.35%~1.80%之间最为合适[53]。还有报道称,在相同的孵化条件下,加入1 μg/L卡马西平可有效提高斑马鱼胚胎的孵化率[54]。谢先中等发现,添加不同浓度的谷氨酰胺(Gln)可以使施氏鲟胚胎发育时间缩短21~8.7 h,孵化率也提高了近11.86%,畸形率显著降低。当浓度超过10 mg/L时,施氏鲟胚胎孵化率下降,畸形率上升;当Gln浓度为5 mg/L时最适合施氏鲟胚胎发育,此时孵化用时最短、孵化率最高,畸形率最低[55]。
在相同的条件下,水流和孵化密度也会影响鱼类胚胎发育。孵化密度过大会造成水体中的溶解氧严重不足,导致鱼卵无法正常发育。水流量小时,影响受精卵与外部物质交换从而影响孵化率;水流量大时可更好地提高水中的溶解氧从而提高孵化率。另外,流水孵化时的畸形率和死亡率远远低于净水孵化[56]。在中华倒刺鲃(Spinibarbus sinensis Bleeker)胚胎发育过程中,水流和孵化密度对胚胎发育均有影响,当水流量大且溶解氧充足时,孵化密度对胚胎发育影响较小;而当水流量和孵化密度的数值比大于1/2时,比较适宜中华倒刺鲃胚胎的发育[57]。
上述研究表明,饲料中的抗生素、水中的污染物、重金属等微量元素均会对鱼类胚胎发育产生不良影响。尤其在自然水域,重金属含量超标会严重影响野生鱼类的正常产卵、发育和生长。目前,我国人工繁殖普遍采用地下水或经过消毒处理的水,水质较好,均可正常进行鱼苗孵化工作。对于一些鱼类,通过配制适量的药物或试剂可以更好地促进鱼类胚胎的正常发育。
3 鱼类杂交胚胎发育研究
一直以来杂交广泛用于农牧业中的品种改良,通过杂交可以结合父母本的基因获得具有杂交优势的后代。而在鱼类杂交育种中,大多数鱼类杂交出现了胚胎无法正常发育、畸形、孵化率低等现象[58]。但也有部分鱼类如异育银鲫[17]、建鲤(Cyprinus carpio var. Jian)[59]、和杂交鲟“鲟龙1号”[60]、牙鲆“鲆优1号”[61]等分别在生长、肉质、抗病及抗寒方面呈现了较好的杂交优势。通过观察研究,杂交鱼类胚胎发育情况对提高其孵化率、成活率以及培育新品种具有意义。田永胜等在蓝身大斑石斑鱼(Epinephelus tukula)(♂)冷冻精子与棕点石斑鱼(E. fuscoguttatus)(♀)杂交试验中发现,其受精率、孵化率和畸形率均无明显差异,当蓝身大斑石斑鱼和鞍带石斑鱼(E. lanceolatus)作为雄鱼与棕点石斑鱼杂交时,蓝身大斑石斑鱼具有更好的杂交优势[62];成美玲等在对鞍带石斑鱼(♀)×蓝身大斑石斑鱼(♂)杂交试验中发现,当水温相同时,杂交组与鞍带石斑鱼(对照组)的胚胎孵化时间差异并不明显,杂交组的受精率、孵化率略微低于对照组,但差异并不明显。另外,在仔鱼阶段的特定时期内杂交组的生长速率显著优于对照组,且杂交组与母本相比有明显生长优势[63]。该试验结果与云纹石斑鱼(♀)×蓝身大斑石斑鱼(♂)杂交的结果相似,当孵化温度控制在(24.8 ± 0.5) ℃时,其胚胎均能正常发育,杂交子代的受精率、孵化率均有略微的下降,畸形率有所上升,其卵经与对照组(云纹石斑鱼)相比略大且孵化时间慢于对照组1.5 h[64],与其他石斑鱼属相比,其孵化时间有较大差异,这也与温度及卵径大小有密切的关系。但在孵化后杂交子代的生长速率明显优于对照组,具有良好的杂交优势。在鞍带石斑鱼(♀)×云纹石斑鱼(♂)杂交育种试验中,当温度为24 ℃时,其胚胎均能正常发育,孵化用时为18 h 52 min,发现正反交子代的发育速率均快于云纹石斑鱼,但慢于鞍带石斑鱼,而正交子代生长速率却快于于反交子代[65],其子代发育速率与杨求华等对云纹石斑鱼(♀)×赤点石斑鱼(E. akaara)(♂)杂交的比较相似[66]。这些研究结果都为石斑鱼杂交优势性状形成及遗传信息提供了一定的基础。此外,杨洋等首次对短须裂腹鱼(Schizothorax wangchiachii)(♀)×鲈鲤(Percocyprispingi pingi)(♂)进行远缘杂交时发现,当孵化温度控制在(14.5±1.0) ℃时,其杂交子一代胚胎受精率高达95.33%,孵化率为71.12%,胚胎发育速率均快于父母本,而这也可能与胚胎在胚孔封闭期有轻微的卵黄运动有关,但在仔鱼期杂交子一代生长速度慢于父本鲈鲤;而在反交试验中,其杂交子代胚胎发育至原肠晚期时死亡率超过85%,最后仅有的12尾出膜后都出现畸形现象并在3 d后死亡[67]。施氏鲟(Acipenser schrenckii)(♀)×西伯利亚鲟(♂)(A. baeri)杂交子代的受精率最高达85.6%,由于受到孵化温差较大原因,其孵化率仅为55%,远低于父母本的正常孵化率,所以把温度在一定温度范围内还可以进一步提高杂交子代的孵化率[68]。在达氏鳇(♀) ×施氏鲟(♂)杂交的过程中,无论正交还是反交,其孵化率和受精率差异并不显著,均超过81%,出苗率也超过79%,与施氏鲟差异均不显著[69]。在观赏鱼中,通过对红头丽体鱼(Cichlasoma synspilum)(♀)×红魔丽体鱼(C. citrinellum)(♂)进行杂交,其胚胎在水温为(30±05) ℃的情况下,孵化时间为 52 h 36 min,受精率与孵化率均超过91%,均高于其他杂交鱼类的受精率和孵化率,表明这2种鱼有较强的杂交亲和性[70]。通过花(Hemibarbus maculatus)(♀)×唇(H. labeo)(♂)杂交发现,其子代胚胎发育时间与母本花胚胎发育时间比较相近,与父本唇胚胎发育相比有较大的差异[71]。该试验结果与相关报道非常相似,如翘嘴鳜(Siniperca chuatsi)(♀)×斑鳜(S. scherzeri)(♂)[72]、状黄姑鱼(Nibea miichthioides)(♀)×大黄鱼(Pseudosciaena crocea)(♂)[73]、星斑川鲽(Platichthys stellatus)和石鲽(Kareius bicoloratus)[74]等鱼类杂交试验,均发现其子代胚胎发育时间与母本胚胎发育时间比较一致。上述研究结果表明,在相同的条件下,大多数鱼类在杂交育种过程中都有一定的杂交优势。其中,杂交子代的胚胎发育过程、时间均与母本比较接近,并有一定的杂交优势,与父本的胚胎发育过程有较大差异。这些杂交鱼类胚胎发育特点表明,杂交鱼类中母本雌核基因可能在杂交胚胎发育过程中发挥了一定的作用(表2)。
4 展望
近几年,水资源污染对我国渔业资源造成了严重的威胁,尤其在鱼类产卵期,严重影响我国相关鱼类胚胎的正常发育和生长,造成野生鱼类资源锐减、种质资源退化等现象。相关部门应加大监管管理力度,避免水中的重金属元素等有害污染物对鱼类胚胎发育及后期的生长造成不良影响。
鱼类胚胎发育观察研究一直都是一个相对传统但又非常重要的的研究热点。近些年,学者们对各种鱼类进行了胚胎发育观察,越来越多的鱼类被开发、保存下来,但大多数研究主要集中在胚胎发育观察和温度对整个胚胎发育速率、受精率等方面,仍不够系统、深入。随着生物技术以及相关智能设备的发展和完善,还可以对鱼类胚胎进行更深入、系统的研究,如:(1)深入研究添加中草药、植物等提取物对胚胎发育的影响,有学者发现,从柚子皮中提取的果胶可以作为鱼类天然的孵化剂,通过上调斑马鱼孵化酶(ZHE1)来促进斑马鱼孵化能力,经果胶(100 μg/mL)处理过的斑马鱼的孵化率(96.6%)显著高于未处理的斑马鱼胚胎的孵化率(66.6%)[75],与化学试剂相比,通过开发研究天然孵化剂对鱼类胚胎发育的影响,将会更有利于鱼类胚胎发育及水环境的保护;(2)原肠期是鱼类胚胎发育阶段对温度变化的“最敏感时期”,其他发育阶段对温度变化等相关因素是否存在“敏感期”,其作用机制又是什么,可以根据这些去探讨不同温度、盐度以及其他环境因子对胚胎不同发育阶段的影响等方面的研究;(3)如何提高鱼类胚胎的免疫保护力以及相关鱼类对外界环境因素的免疫力是否相同等问题。目前,针对这些问题还需进一步地探究。未来随着鱼类胚胎发育研究的不断深入以及上述问题的解决,在增产增效的同时,还将对我国鱼类发育学、鱼类繁育、濒危鱼类保护、鱼类进化和渔业资源又好又快发展具有重要意义。
参考文献:
[1]Herzing A,Winkler H. The influence of temperature on the embryonic development of three cyprinid fishes,Abramis brama,Chalcalburnus chalcoides mento and Vimba vimba[J]. Journal of Fish Biology,1986,28(2):171-181.
[2]殷名称. 鱼类生态学[M]. 北京:中国农业出版社,1995:105-130.
[3]郑金霞,范兆廷. 虹鳟胚胎受精率适宜发育期的研究[J]. 水产学杂志,2012,25(2):31-34.
[4]柴学军,孙 敏,许源剑. 温度和盐度对日本黄姑鱼胚胎发育的影响[J]. 南方水产科学,2011,7(5):43-49.
[5]何华敏. 乌苏里拟鲿养殖生物学研究[D]. 苏州:苏州大学,2011.
[6]乔德亮,付立霞. 斑点叉尾胚胎及卵黄囊期仔鱼发育研究[J]. 水生态学杂志,2009,30(1):58-63.
[7]黄小林,李 涛,林黑着,等. 网箱养殖黄斑篮子鱼胚胎发育观察[J]. 南方水产科学,2018,14(2):96-101.
[8]张燕萍,章海鑫,傅义龙,等. 黄尾鲴的胚胎发育[J]. 江苏农业科学,2019,47(16):174-178.
[9]Bagenal T B,Braum E. Eggs and early life history of fish [J]. Ibp Handbook,1978:165-201.
[10]董 纯,陈小娟,万成炎,等. 圆口铜鱼人工繁殖及胚胎发育研究[J]. 水生态学杂志,2019,40(3):115-119.
[11]时红卫,吴美琴,张俊彬. 鲻鱼人工繁殖与胚胎发育观察[J]. 上海海洋大学学报,2015,24(6):855-861.
[12]万 远,占 阳,欧阳珊,等. 胭脂鱼胚胎及仔鱼早期发育观察[J]. 南昌大學学报(理科版),2013,37(1):78-82.
[13]杜 伟,蒙子宁,薛志勇,等. 半滑舌鳎胚胎发育及其与水温的关系[J]. 中国水产科学,2004,11(1):48-53.
[14]郭长江,赵 文,石振广,等. 达氏鳇养殖群体的胚胎发育研究[J]. 大连海洋大学学报,2016,31(6):589-597.
[15]Bai L Y,Zhao Z Y. Study on ebryonicdvelopment,lrva,jvenile and yungfsh of Epinephelus fuscoguttatus[J]. Agriculture science & Technology,2016,17(6):1305-1314.
[16]林红军,刘 莎,曾 玲. 黄颡鱼人工授精及胚胎发育研究[J]. 黑龙江畜牧兽医,2019(12):138-140,170.
[17]薛凌展. 温度对异育银鲫中科3号胚胎发育的影响[J]. 福建师范大学学报(自然科学版),2014,30(4):76-83.
[18]骆小年,段友健,郭 童,等. 北方须鳅人工繁殖与胚胎发育研究[J]. 大连海洋大学学报,2021(2):187-194.
[19]陈福艳,梁万文,冯鹏霏,等. 丁鱼胚胎及仔鱼发育的观察[J]. 南方农业学报,2012,42(3):315-319.
[20]郭永灿. 水温对鲢鱼、草鱼胚胎发育的影响[J]. 淡水渔业,1982(3):35-40.
[21]刘艳超,刘海平,刘书蕴,等. 温度对尖裸鲤胚胎发育及其仔稚鱼生长性状的影响[J]. 动物学杂志,2018,53(6):910-923.
[22]王亚利,解崇友,尹 敏,等. 温度对四川华鳊胚胎发育的影响[J]. 水产科学,2020,39(1):79-87.
[23]袁宗勤,于燕光,叢日浩,等. 温度对斑马鱼胚胎发育的影响研究[J]. 现代农业科学,2009(4):188-189,220.
[24]黄贤克,单乐州,闫茂仓,等. 黄姑鱼胚胎发育及其与温度和盐度的关系[J]. 海洋科学,2017,41(7):44-50.
[25]黄永春,胡石柳,周泽斌,等. 状黄姑鱼早期发育特征及对温盐度的适应性[J]. 中国海洋大学学报(自然科学版),2006,36(1):101-106.
[26]史秀兰,王炳谦,黄天晴,等. 黑龙江乌苏里白鲑(Coregonus ussuriensis Berg)胚胎发育观察研究[J]. 海洋与湖沼,2020,51(2):415-421.
[27]崔文耀,周丽青,杨继辉,等. 四带小鲃胚胎发育及系统进化分析[J]. 水产学报,2020,44(8):1286-1295.
[28]Wang F,Wang X G,Liu C,et al. Transparent tiger barb Puntius tetrazona,a fish model for in vivo analysis of nocardial infection[J]. Veterinary Microbiology,2017,211:67-73.
[29]王友红,刘洪军,于道德,等. 海水青鳉胚胎发育的观察[J]. 海洋科学,2017,41(6):18-25.
[30]陈 漪,王晓杰,冉皓宇,等. 海水模式种青鳉鱼(Oryzias melastigma)的胚胎发育观察[J]. 海洋与湖沼,2016,47(1):71-82.
[31]刘海平,刘孟君,牟振波,等. 西藏双须叶须鱼早期发育特征[J]. 水生生物学报,2019,43(5):1041-1055.
[32]官曙光,刘洪军,李祥东,等. 黑棘鲷胚胎发育过程及特殊结构观察[J]. 海洋科学,2011,35(9):68-72.
[33]Hotta K,Tamura M,Watanabe T,et al. Changes in spawning characteristics of Japanese whiting Sillago japonica under control of temperature[J]. Fisheries scienence,2001,67(6):1111-1118.
[34]尹家胜,沈俊宝,徐 伟,等. 水温变化对绥芬河滩头雅罗鱼产卵的影响[J]. 动物学报,2001,47(6):704-708.
[35]吴利敏,袁淑仪,赵晨华,等. 不同温度对淇河鲫胚胎发育的影响[J]. 山西农业大学学报(自然科学版),2018,38(7):50-56.
[36]施兆鸿,夏连军,王建钢,等. 盐度对黄鲷胚胎发育及早期仔鱼生长的影响[J]. 水产学报,2004,28(5):599-602.
[37]刘伟成,冀德伟,单乐州,等. 温度和盐度对条石鲷胚胎发育的影响[J]. 水生态学杂志,2010,3(6):101-104.
[38]蔡文超,区又君,李加儿. 盐度对条石鲷胚胎发育的影响[J]. 生态学杂志,2010,29(5):951-956.
[39]宋振鑫,陈 超,吴雷明,等. 盐度与pH对云纹石斑鱼胚胎发育和仔鱼活力的影响[J]. 渔业科学进展,2013,34(6):52-58.
[40]赵 明,陈 超,柳学周,等. 盐度对七带石斑鱼胚胎发育和卵黄囊仔鱼生长的影响[J]. 渔业科学进展,2011,32(2):16-21.
[41]张培军. 海水鱼类系列发育和养殖生物学[M]. 济南:山东科学技术出版社,1999:104-110.
[42]史晋绒,王永明. Cu2+对稀有鲫胚胎发育的影响[J]. 内江师范学院学报,2014,29(6):47-51.
[43]刘 丽. CuInS2/ZnS QDs对稀有鲫的生殖及胚胎发育毒性效应研究[D]. 重庆:西南大学,2017:13-27.
[44]Speranza A W,Seeley R J,Seeley V A,et al. The effect of sublethal concentrations of zinc on reproduction in the zebrafish,Brachydaniorerio Hamilton-Buchanan[J]. Environmental Pollution,1977,12(3):217-222.
[45]黄 伟. 汞、铅、锌对褐牙鲆(Paralichthys olivaceus)早期发育过程毒理作用的研究[D]. 青岛:中国科学院研究生院(海洋研究所),2010:35-58.
[46]叶辽辽,徐 胜,袁秋芸,等. SDBS对大鳞副泥鳅胚胎发育及幼鱼性腺发育的影响[J]. 淡水渔业,2018,48(1):86-90.
[47]Wang J,Li J J,Xu N,et al. Responses of Takifugu obscurus fertilized eggs and larvae to increased ammonia exposure[J]. Environmental Science and Pollution Research International,2015,22(20):15976-15984.
[48]Chen Y F,Li J J,Sun H J,et al. Dose-dependent effect of un-ionized ammonia on hatching success of fertilized eggs of silver carp Hypophthalmichthys molitrix and bighead carp Hypophthalmichthys nobilis[J]. Journal of Freshwater Ecology,2013,28(1):39-46.
[49]王梦杰,王海华,马本贺,等. 氨氮对台湾泥鳅胚胎及卵黄囊期仔鱼的毒理效应[J]. 水产科学,2020,39(4):483-490.
[50]王能汉,邓巧玲,范垚城,等. 长江岸线宜昌江段五种不同类型水样对稀有鲫胚胎的毒性[J]. 水生生物学报,2020,44(5):1064-1069.
[51]刘丽丽,吕 鹏,闫艳春. 磺胺二甲嘧啶对斑马鱼胚胎的急性毒性作用[J]. 中国渔业质量与标准,2018,8(1):34-39.
[52]齐澈力木格,尹晓宇,李嘉伟,等. 母体暴露环境浓度盐酸四环素对F1代斑马鱼胚胎骨骼发育的影响[J/OL]. 生态毒理学报,2021,16(2):235-244.
[53]李爱君,张玉茹,夏雪梅,等. 乙醇对斑马鱼胚胎发育的急性毒害效应[J]. 西华大学学报(自然科学版),2019,38(6):27-31.
[54]Qiang L,Cheng J,Yi J,et al. Environmental concentration of carbamazepine accelerates fish embryonic development and disturbs larvae behavior[J]. Ecotoxicology,2016,25(7):1426-1437.
[55]谢先中,陈晓武,冉光鑫,等. 添加谷氨酰胺对施氏鲟胚胎发育及各发育期抗氧化生化指标的影响[J]. 水产学杂志,2018,31(5):10-13.
[56]陈永柏,廖文根,彭期冬,等. 四大家鱼产卵水文水动力特性研究综述[J]. 水生态学杂志,2009,2(2):130-133.
[57]李卿青,任名栋,朱庭耀,等. 孵化密度和水流量对中华倒刺鲃胚胎发育的影响[J]. 水产科技情报,2019,46(1):19-23.
[58]Arai K. Developmental genetic studies on salmonids:morphogenesis,isozyme phenotypes and chromosomes in hybrid embryos[J]. Mem Fac Fish Hokkaido Univ,1984,31:1-94.
[59]張建森,孙小异. 建鲤新品系的选育[J]. 水产学报,2007,31(3):287-292.
[60]张 颖,孙大江,王 斌,等. 杂交鲟鲟龙1号[J]. 中国水产,2017(5):62-67.
[61]田永胜,齐文山,姜 静,等. 鲆优1号牙鲆和亲本群体的表型及遗传性状变异分析[J]. 海洋学报(中文版),2014,36(6):75-86.
[62]田永胜,唐 江,马文辉,等. 蓝身大斑石斑鱼冷冻精子(♂)与棕点石斑鱼(♀)杂交后代发育及生长特征分析[J]. 渔业科学进展,2019,40(6):36-47.
[63]成美玲,田永胜,吴玉萍,等. 鞍带石斑鱼(♀)×蓝身大斑石斑鱼(♂)杂交F1代变态发育和生长特征分析[J]. 海洋学报,2019,41(8):52-62.
[64]唐 江,田永胜,李振通,等. 云纹石斑鱼(♀)×蓝身大斑石斑鱼(♂)远缘杂交后代 发育与生长[J]. 广东海洋大学学报,2019,39(4):27-34.
[65]李子奇,成美玲,吴玉萍,等. 鞍带石斑鱼(♀)×云纹石斑鱼(♂)杂交后代早期发育及正反交后代生长特性[J]. 水产学报,2020,44(3):436-446.
[66]杨求华,黄种持,郑乐云,等. 云纹石斑鱼(♀)×赤点石斑鱼(♂)杂交子代胚胎发育及生长[J]. 海洋渔业,2014,36(3):224-231.
[67]杨 洋,陈 瑶,万玉芳,等. 短须裂腹鱼与鲈鲤杂交F1代胚胎及仔稚鱼发育观察[J]. 南方水产科学,2018,14(6):66-73.
[68]付佳琪,聂竹兰,罗秀华,等. 施氏鲟Acipenser schrenckii♀×西伯利亚鲟A. baeri♂杂交子代胚胎发育的观察[J]. 水产学杂志,2017,30(5):18-22.
[69]张 颖,徐 伟,孙大江,等. 达氏鳇、施氏鲟及其杂交子代胚胎发育期的抗氧化酶活性[J]. 水产学杂志,2017,30(6):19-23.
[70]孙志景,姜巨峰,傅志茹,等. 红头丽体鱼×红魔丽体鱼杂交子一代胚胎发育及仔鱼形态学观察[J]. 南方水产科学,2014,10(3):38-46.
[71]练青平,宓国强,胡廷尖,等. 花(♀)×唇(♂)杂交子一代胚胎发育研究[J]. 湖北农业科学,2012,51(1):131-135.
[72]宓国强,练青平,王雨辰,等. 翘嘴鳜(♀)×斑鳜(♂)杂交子一代的胚胎发育[J]. 上海海洋大学学报,2009,18(4):421-427.
[73]马 梁,王 军,陈武各,等. 状黄姑鱼与大黄鱼人工杂交子代的胚胎发育[J]. 厦门大学学报(自然科学版),2002,41(3):378-382.
[74]马贵范,刘振华,田永胜,等. 星斑川鲽(♀)与石鲽(♂)杂交子代胚胎发育研究[J]. 水产科学,2018,37(2):167-173.
[75]Dananjaya S S,Chandrarathna H U,Nayanaransi L U,et al. Pectin purified from pomelo (Citrus maxima) peel as a natural hatching agent for fish embryos[J]. Aquaculture Research,2020,51(8):3109-3118.
