LncRNA DANCR靶向miR-423-5p调控ox-LDL诱导的血管内皮细胞凋亡、炎症反应①
2021-09-25李骞李秀研林丽丽许景芬陈芬
李骞 李秀研 林丽丽 许景芬 陈芬
(福建医科大学附属泉州第一医院全科医学科,泉州362000)
动脉粥样硬化(atherosclerosis,AS)是一种由多种危险因素引起的慢性炎症性血管疾病,是全球范围内死亡率和发病率升高的主要原因[1]。内皮细胞在血管功能中发挥重要作用,其功能障碍被认为是AS发生的重要标志。氧化低密度脂蛋白(oxidized low-density lipoprotein,ox-LDL)是诱导内皮细胞损伤的关键因子,可刺激内皮细胞炎症反应和氧化应激,增加细胞凋亡,破坏细胞功能,进而促进AS发生发展[2]。因此,抑制ox-LDL诱导的内皮细胞损伤对防治AS意义重大。长链非编码RNA(long non-cod‐ing RNA,LncRNA)是一类长度超过200个核苷酸的非编码RNA,其通过在转录后、转录和染色质水平上调控基因表达,参与调控内皮细胞功能障碍、胆固醇积聚、炎症等病理过程,与AS进展密切相关[3]。分化拮抗非蛋白编码RNA(differentiation antagonis‐tic non-protein coding RNA,DANCR)是一种肿瘤相关lncRNA,包括肝癌、乳腺癌和肺癌在内的多种恶性肿瘤中DANCR表达失调,其异常表达有助于癌细胞的增殖、迁移和侵袭,是一种潜在的癌症标志物和治疗靶点[4-5]。此外,DANCR还可提高肾小管上皮细胞活力,抑制脂多糖诱导的炎症反应和凋亡[6]。然而,XIST介导AS进展的潜在机制仍不清楚。本研究采用ox-LDL诱导人脐静脉内皮细胞(human umbilical vein endothelial cell,HUVEC)模拟AS细胞损伤,重点探讨LncRNA DANCR对ox-LDL诱导的HUVEC凋亡和炎症因子分泌的作用,分析其潜在靶基因,以期为AS治疗提供新的方向。
1 材料与方法
1.1 材料HUVEC购于中国典型培养物保藏中心;DMEM培养基、胎牛血清购于美国Sigma公司;一步法PCR逆转录试剂盒、微小RNA(microRNA,miRNA)逆转录试剂盒、SYBR Green master Mix购于北京天根生化科技有限公司;miRNA模拟物(miR‐NA mimics)、miRNA抑制物(anti-miR)、过表达载体(pcDNA-RNA)、小干扰RNA(si-RNA)、双荧光素酶报告基因载体由上海生工公司提供;ox-LDL、Trizol试剂、细胞计数试剂盒(cell counting kit 8,CCK-8)、膜联蛋白V-异硫氰酸荧光素/碘化丙啶(Annexin VFITC/PI)凋亡检测试剂盒购于北京索莱宝生物科技公司;细胞周期素D1(CyclinD1)小鼠单克隆抗体、活化型半胱氨酸蛋白酶3(Cleaved-caspase-3)兔单克隆抗体、β-actin兔单克隆抗体、羊抗兔IgG、羊抗鼠IgG购于上海碧云天生物公司;IL-1β、IL-6、TNF-α ELISA检测试剂盒购于上海冠导生物工程有限公司。
1.2 方法
1.2.1 HUVEC体外损伤模型建立用含10%胎牛血清的DMEM培养基置于含5%CO2、37℃恒温细胞培养箱中培养HUVEC,然后用含50 mg/L ox-LDL的细胞培养液处理细胞24 h构建体外损伤模型,模拟HUVEC AS损伤,记为OX-LDL组,同时设置正常对照(normal control,NC)组[7]。
1.2.2 细胞转染和实验分组将对数期HUVEC接种于6孔板,利用脂质体转染法分别将pcDNA-con、pcDNA-DANCR、anti-miR-con、anti-miR-423-5p、pcD‐NA-DANCR+miR-con、pcDNA-DANCR+miR-423-5p mimics分别转染HUVEC,转染成功后用含50 mg/L ox-LDL的细胞培养液处理细胞24 h,依次命名为ox-LDL+pcDNA-con组、ox-LDL+pcDNA-DANCR组、ox-LDL+anti-miR-con组、ox-LDL+anti-miR-423-5p组、ox-LDL+pcDNA-DANCR+miR-con组、ox-LDL+pcDNADANCR+miR-423-5p组。
1.2.3 实时荧光定量PCR(RT-qPCR)检测Ln‐cRNA DANCR和miR-423-5p表达按照Trizol试剂说明书提取细胞总RNA,测定浓度和纯度后-20℃保存备用。取适量的总RNA,分别使用一步法PCR逆转录试剂盒或miRNA逆转录试剂盒合成cDNA,用SYBR Green master Mix进行RT-qPCR扩增。具体引物如下(5'-3'):DANCR上游引物CTGCATTCCTGAACCGTTATCT,DANCR下游引物GGGTG‐TAATCCACGTTTCTCAT;β-actin上游引物CCAACCGCGAGAAGATGA,β-actin下游引物CCAGAGGC‐GTACAGGGATAG;miR-423-5p上游引物ATGGTTC‐GTGGGTGA,miR-423-5p下 游 引 物GTGCAGGGTCCGAGGT;U6上游引物CTCGCTTCGGCAGCACA,U6下游引物AACGCTTCACGAATTTGCGT。2-ΔΔCt法计算LncRNA DANCR(内参为β-actin)和miR-423-5p(内参为U6)相对表达量。
1.2.4 CCK-8法检测细胞活力收集各组细胞,按照100 μl/孔、5×103个/孔接种于96孔板,24 h后每孔加入10 μl的CCK-8溶液,培养箱继续孵育2.5 h,酶标仪检测450 nm波长处每孔的吸光度(A)值。细胞存活率(%)=A实验组/A对照组×100%。
1.2.5 流式细胞术检测细胞凋亡收集各组细胞,结合缓冲液调整为1×108个/L的单细胞悬液。取100 μl细胞悬液,按照Annexin V-FITC/PI双染法细胞凋亡检测试剂盒分别加入Annexin V-FITC和PI,室温条件下暗室孵育15 min,补加结合缓冲液至总体积为500 μl,流式细胞术检测细胞凋亡。
1.2.6 Western blot检 测CyclinD1、Cleaved-cas‐pase-3蛋白表达水平放射免疫沉淀缓冲液提取各组细胞的总蛋白。蛋白定量后取适量蛋白和等体积上样缓冲液混合,煮沸5 min使蛋白变性冷却至室温后备用。按照每泳道30 μg蛋白上样,100 V 90 min进行聚丙烯酰胺凝胶电泳。快速蛋白湿转移将蛋白转移至硝酸纤维素膜后,用含5%脱脂奶粉的封闭液室温孵育2 h。用稀释的CyclinD1抗体(1∶200)、Cleaved-caspase-3抗体(1∶1 000)、β-actin抗体(1∶1 000)溶液孵育室温孵育膜2 h。用稀释的羊抗兔IgG、羊抗鼠IgG溶液(1∶1 000)室温孵育膜1 h。用化学发光显色试剂盒进行显影。曝光后,Image J1.8.0软件检测每个条带灰度值,以目的蛋白与内参蛋白β-actin灰度值比值表示目的蛋白相对表达水平。
1.2.7 双荧光素酶报告基因实验将含有miR-423-5p结合位点的LncRNA DANCR野生(WT)序列或突变(MUT)序列克隆到双荧光素酶报告质粒,构建双荧光素酶报告基因载体WT-LncRNA DANCR、MUT-LncRNA DANCR。利用脂质体转染法将WTLncRNA DANCR、MUT-LncRNA DANCR分 别 与miR-con或miR-423-5p mimics共 转 染 至HUVEC,48 h后检测细胞相对荧光素酶活性。同时将pc-DNA-con、pcDNA-DANCR、si-con、si-DANCR分别转染HUVEC,48 h后RT-qPCR检测miR-423-5p表达水平。
1.2.8 ELISA法检测IL-1β、IL-6、TNF-α分 泌水平收集各组细胞上清液,参照ELISA试剂盒说明书步骤分别检测IL-1β、IL-6、TNF-α水平。
1.3 统计学分析每组设置3个平行实验,重复3次,所有计量资料符合正态分布,用±s表示。采用SPSS22.0软件进行统计学分析,两组间比较采用t检验,多组间比较采用单因素方差分析,组件多重比较采用SNK-q检验。P<0.05为差异具有统计学意义。
2 结果
2.1 ox-LDL处理HUVEC中LncRNA DANCR、miR-423-5p的表达情况与NC组比较,ox-LDL组HU‐VEC中LncRNA DANCR的表达水平显著降低,miR-423-5p表达水平显著升高(P<0.05)。见表1。
表1 HUVEC中LncRNA DANCR和miR-423-5p表 达 量(±s,n=9)Tab.1 Expressions of LncRNA DANCR and miR-423-5p in HUVEC(±s,n=9)

表1 HUVEC中LncRNA DANCR和miR-423-5p表 达 量(±s,n=9)Tab.1 Expressions of LncRNA DANCR and miR-423-5p in HUVEC(±s,n=9)
Note:Compared with NC,1)P<0.05.
?
2.2 过表达DANCR对ox-LDL诱导的HUVEC增殖和凋亡的影响与NC组比较,ox-LDL组HUVEC中LncRNA DANCR表达显著降低,细胞存活率、Cy‐clinD1蛋白表达显著降低,细胞凋亡率、Cleaved-cas‐pase-3蛋白表达显著升高(P<0.05);与ox-LDL+pcDNA-con组比较,ox-LDL+pcDNA-DANCR组HU‐VEC中LncRNA DANCR表达显著升高,细胞存活率、CyclinD1蛋白表达显著升高,细胞凋亡率、Cleaved-caspase-3蛋白表达显著降低(P<0.05)。见图1、表2。
表2 过表达DANCR对ox-LDL诱导的HUVEC细胞增殖和凋亡的影响(±s,n=9)Tab.2 Effect of DANCR overexpression on proliferation and apoptosis of ox-LDL-induced HUVEC cells(±s,n=9)
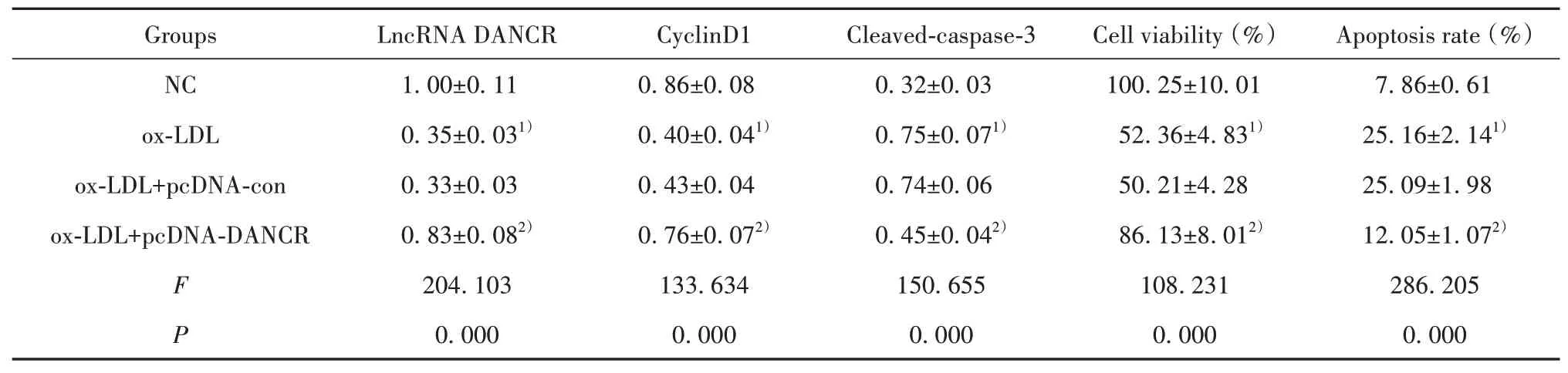
表2 过表达DANCR对ox-LDL诱导的HUVEC细胞增殖和凋亡的影响(±s,n=9)Tab.2 Effect of DANCR overexpression on proliferation and apoptosis of ox-LDL-induced HUVEC cells(±s,n=9)
Note:Compared with NC,1)P<0.05;compared with ox-LDL+pcDNA-con,2)P<0.05.
?
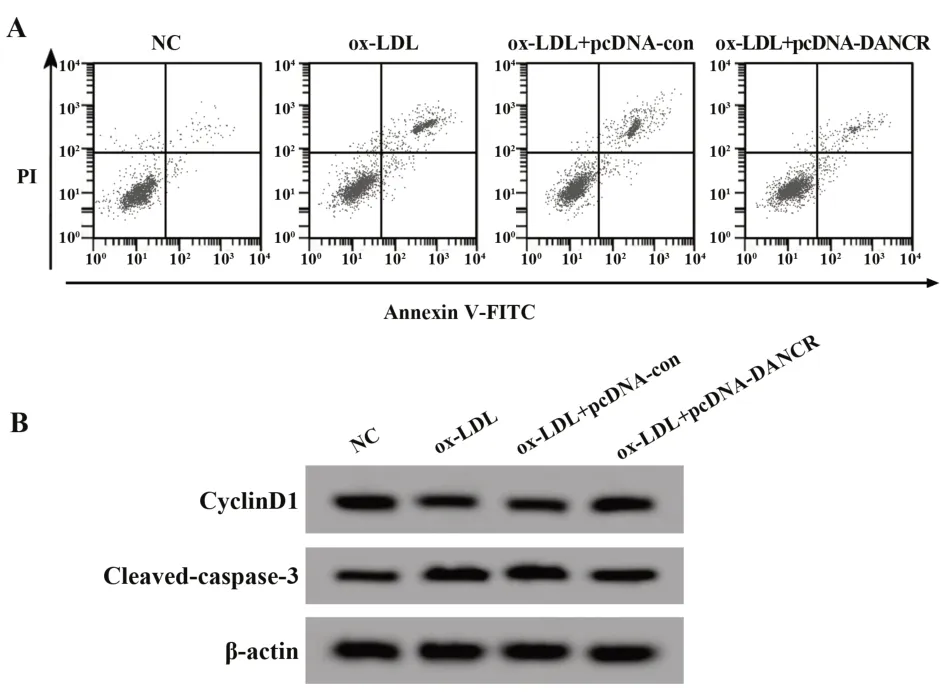
图1 过表达DANCR对ox-LDL诱导的HUVEC增殖和凋亡的影响Fig.1 Effect of DANCR overexpression on proliferation and apoptosis of ox-LDL-induced HUVEC
2.3 抑 制miR-423-5p对ox-LDL诱导 的HUVEC增殖和凋亡的影响与ox-LDL+anti-miR-con组比较,ox-LDL+anti-miR-423-5p组HUVEC中miR-423-5p表达显著降低,细胞存活率、CyclinD1蛋白表达显著升高,细胞凋亡率、Cleaved-caspase-3蛋白表达显著降低(P<0.05)。见表3、图2。
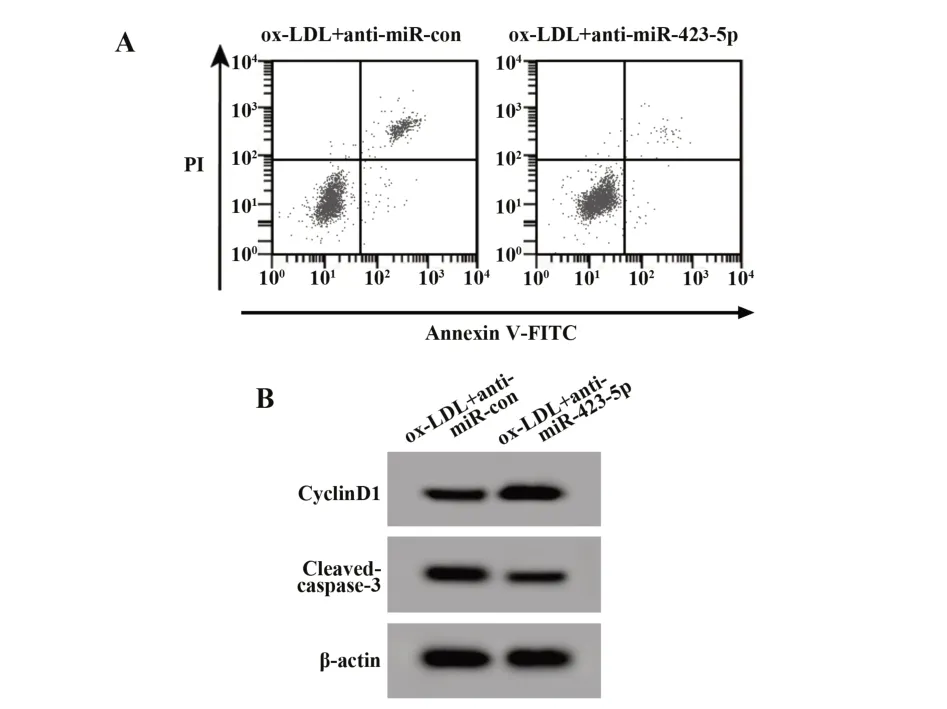
图2 抑制miR-423-5p表达对ox-LDL诱导的HUVEC细胞增殖和凋亡的影响Fig.2 Effect of miR-423-5p inhibition on proliferation and apoptosis of ox-LDL-induced HUVEC cells
表3 抑制miR-423-5p表达对ox-LDL诱导的HUVEC增殖和凋亡的影响(±s,n=9)Tab.3 Effect of miR-423-5p inhibition on proliferation and apoptosis of ox-LDL-induced HUVEC(±s,n=9)
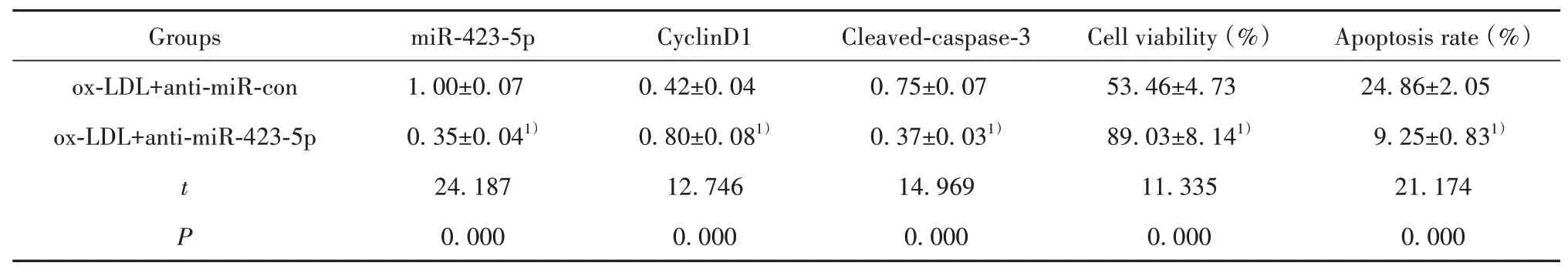
表3 抑制miR-423-5p表达对ox-LDL诱导的HUVEC增殖和凋亡的影响(±s,n=9)Tab.3 Effect of miR-423-5p inhibition on proliferation and apoptosis of ox-LDL-induced HUVEC(±s,n=9)
Note:Compared with ox-LDL+anti-miR-con,1)P<0.05.
?
2.4 LncRNA DANCR靶向miR-423-5p LncRNA DANCR与miR-423-5p之间存在特异性互补结合位点,见图3。与miR-con和WT-LncRNA DANCR共转染比较,miR-423-5p mimics和WT-LncRNA DANCR共转染后HUVEC相对荧光素酶活性显著降低(P<0.05);与miR-con和MUT-LncRNA DANCR共转染比较,miR-423-5p mimics和MUT-LncRNA DANCR共转染后HUVEC相对荧光素酶活性无显著变化,见表4。pcDNA-DANCR组HUVEC中miR-423-5p表达显著低于pcDNA-con组(P<0.05);si-DANCR组HUVEC中miR-423-5p表达显著高于si-con组(P<0.05),见表5。
表4 双荧光素酶报告实验(±s,n=9)Tab.4 Double luciferase report experiment(±s,n=9)
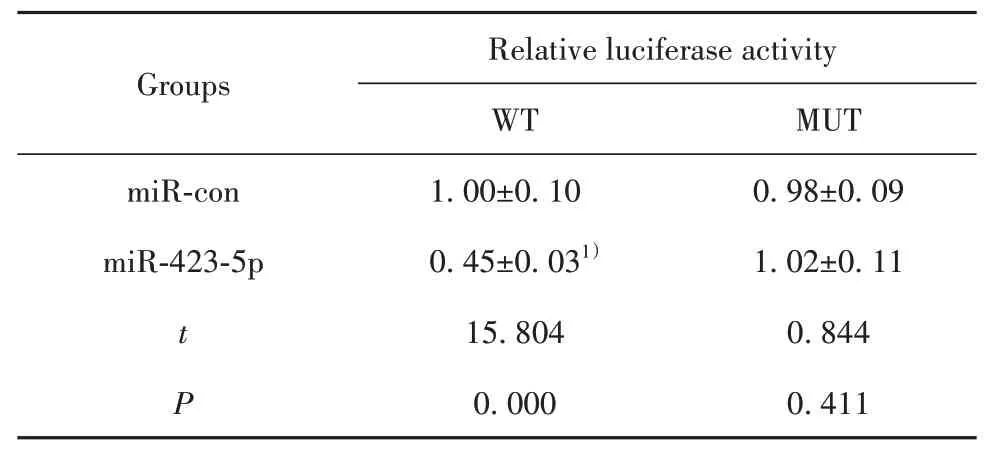
表4 双荧光素酶报告实验(±s,n=9)Tab.4 Double luciferase report experiment(±s,n=9)
Note:Compared with miR-con,1)P<0.05.
?
表5 RT-qPCR检测miR-423-5p的表达(±s)Tab.5 RT-qPCR detects expressions of miR-423-5p(±s)
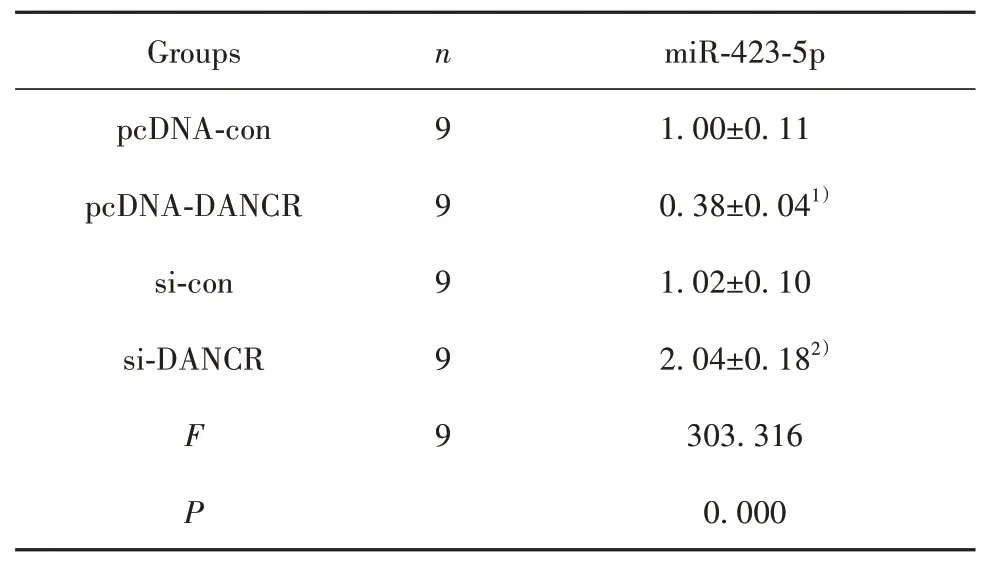
表5 RT-qPCR检测miR-423-5p的表达(±s)Tab.5 RT-qPCR detects expressions of miR-423-5p(±s)
Note:Compared with pcDNA-con,1)P<0.05;compared with si-con,2)P<0.05.
?

图3 miR-423-5p和LncRNA DANCR结合预测示意图Fig.3 Schematic diagram of miR-423-5p and LncRNA DANCR binding prediction
2.5 过表达miR-423-5p可以逆转DANCR过表达对ox-LDL诱导的HUVEC增殖和凋亡的影响与ox-LDL+pcDNA-DANCR+miR-con组 比较,ox-LDL+pcDNA-DANCR+miR-423-5p组HUVEC中miR-423-5p表达显著升高,细胞存活率、CyclinD1蛋白表达显著降低,细胞凋亡率、Cleaved-caspase-3蛋白表达显著升高(P<0.05)。见表6、图4。
表6 过表达miR-423-5p可以逆转DANCR过表达对ox-LDL诱导的HUVEC增殖和凋亡的影响(±s,n=9)Tab.6 miR-423-5p overexpression can reverse effect of DANCR overexpression on proliferation and apoptosis of ox-LDLinduced HUVEC(±s,n=9)
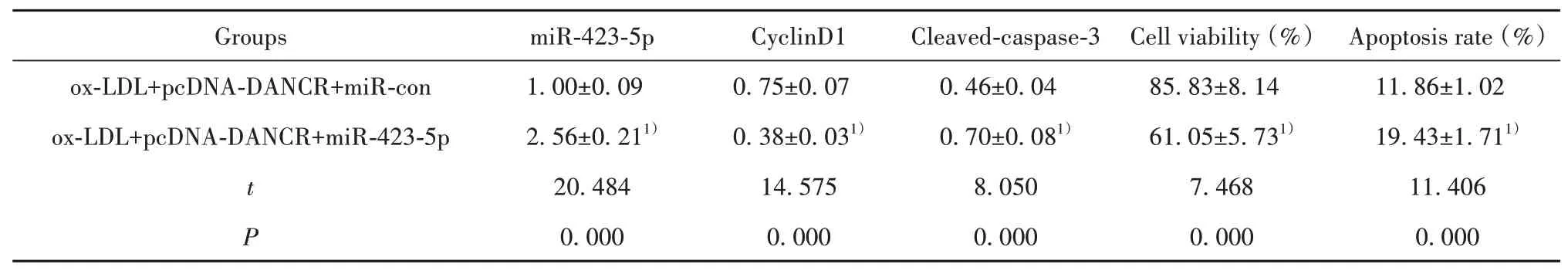
表6 过表达miR-423-5p可以逆转DANCR过表达对ox-LDL诱导的HUVEC增殖和凋亡的影响(±s,n=9)Tab.6 miR-423-5p overexpression can reverse effect of DANCR overexpression on proliferation and apoptosis of ox-LDLinduced HUVEC(±s,n=9)
Note:Compared with ox-LDL+pcDNA-DANCR+miR-con,1)P<0.05.
?
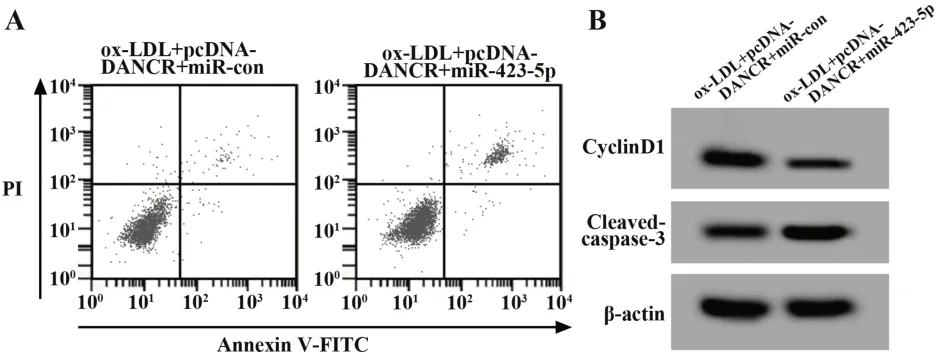
图4 过表达miR-423-5p可以逆转DANCR过表达对ox-LDL诱导的HUVEC细胞增殖和凋亡的影响Fig.4 miR-423-5p overexpression can reverse effect of DANCR overexpression on proliferation and apop⁃tosis of ox-LDL-induced HUVEC cells
2.6 ELISA检测各组炎症因子分泌水平与NC组比较,ox-LDL组HUVEC上清中IL-1β、IL-6、TNF-α水平显著升高(P<0.05);与ox-LDL组比较,ox-LDL+pcDNA-DANCR组、ox-LDL+anti-miR-423-5p组HU‐VEC上清中IL-1β、IL-6、TNF-α水平显著降低(P<0.05);与ox-LDL+pcDNA-DANCR组相比,ox-LDL+pcDNA-DANCR+miR-423-5p组HUVEC上 清 中IL-1β、IL-6、TNF-α水平显著升高(P<0.05)。见表7。
表7 ELISA检测各组炎症因子分泌水平(±s,n=9)Tab.7 ELISA detects secretion levels of inflammatory factors in each group(±s,n=9)
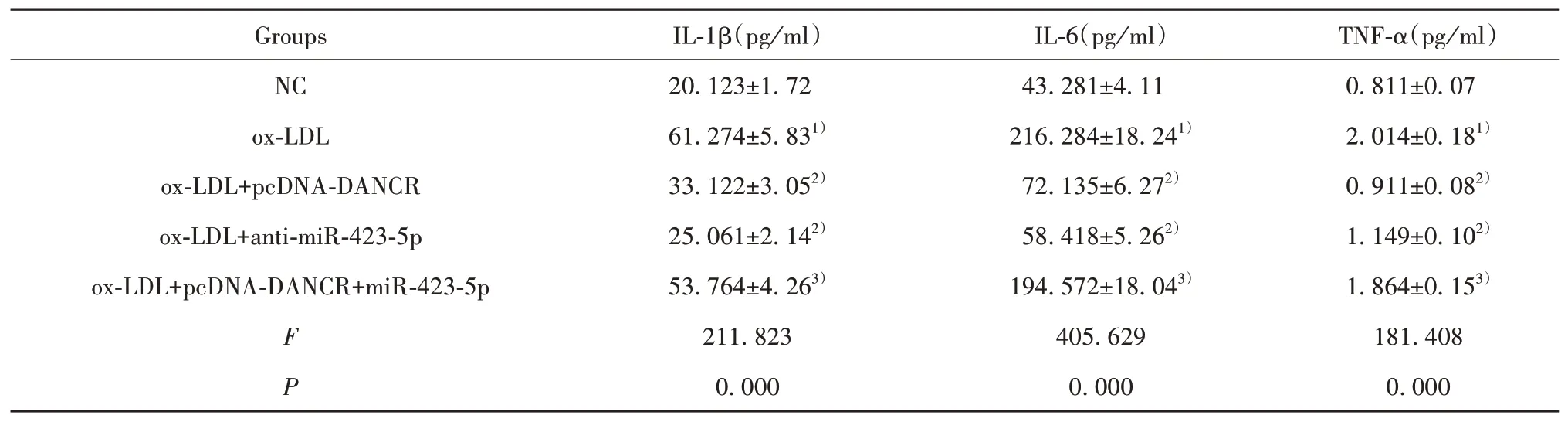
表7 ELISA检测各组炎症因子分泌水平(±s,n=9)Tab.7 ELISA detects secretion levels of inflammatory factors in each group(±s,n=9)
Note:Compared with NC,1)P<0.05;compared with ox-LDL,2)P<0.05;compared with ox-LDL+pcDNA-DANCR,3)P<0.05.
?
3 讨论
AS是冠心病和脑卒中的主要病理基础,是心血管疾病发生的主要原因。内皮细胞损伤及其功能障碍在AS的发展中起着至关重要的作用,探讨与内皮细胞损伤相关的基因,分析其作用机制,有望为AS治疗提供新的策略。
LncRNA DANCR在缺氧诱导的大鼠心肌细胞中表达降低,LncRNA DANCR高表达通过上调缺氧诱导因子1α(hypoxia-inducible factor 1α,HIF-1α)表达减轻缺氧诱导的心肌细胞损伤[8]。LncRNA DANCR高表达可增强糖氧剥夺处理的脑微血管内皮细胞的增殖、迁移和血管生成,对改善缺血性脑卒中具有重要意义[9]。本研究结果表明,ox-LDL处理HU‐VEC后,细胞存活率显著降低,凋亡率显著升高,Lnc-RNA DANCR表达量显著降低。IL-1β、IL-6、TNF-α等炎症因子的分泌是参与AS的发病机制的重要因素[10-11]。本研究显示,ox-LDL诱导后HUVEC上清中IL-1β、IL-6、TNF-α水平显著升高,提示LncRNA DANCR低表达可能与ox-LDL诱导的HUVEC损伤有关。CyclinD1是一种重要的周期蛋白,磷脂酰肌醇激酶抑制剂BYL719通过下调CyclinD1表达可诱导HUVEC阻滞于G1期,抑制细胞增殖[12]。Cas‐pase-3是细胞凋亡发展中最关键的执行分子,其活化是检测细胞凋亡的重要指标[13]。进一步转染DANCR过表达载体上调LncRNA DANCR表达发现,ox-LDL诱导的HUVEC存活率、CyclinD1升高,凋亡率、Cleaved-caspase-3表达降低,IL-1β、IL-6、TNF-α水平降低,说明过表达LncRNA DANCR能够改善ox-LDL诱导的HUVEC炎症反应和凋亡。
miRNA为非编码RNA家族重要成员,研究显示,miRNA通过与LncRNA相互作用参与调解氧化应激、细胞凋亡和炎症反应等多种细胞过程[14-16]。为探索LncRNA DANCR作用机制,本研究发现miR-423-5p是其潜在靶基因。研究表明缺氧诱导的心肌细胞中miR-423-5p表达增加,miR-423-5p高表达导致线粒体膜电位的丢失、活性氧生成增加,促进心肌细胞凋亡[17]。在急性肾损伤后,miR-423-5p通过诱导内质网络应激和氧化应激,抑制肾小管上皮细胞的修复[18]。此外,LncRNA RPSAP52通过靶向下调miR-423-5p能够抑制缺氧诱导的肾近端小管上皮细胞凋亡[19]。本研究显示,ox-LDL诱导后HU‐VEC中miR-423-5p表达显著升高,抑制miR-423-5p表达可提高HUVEC存活率,减轻ox-LDL诱导的HUVEC凋亡、炎症因子分泌,与过表达LncRNA DANCR作用一致。进一步研究证实,LncRNA DANCR对miR-423-5p具有靶向负调控作用,且过表达miR-423-5p能够逆转LncRNA DANCR对ox-LDL处理的HUVEC炎症因子分泌、存活、凋亡以及相关蛋白表达的影响,说明LncRNA DANCR可提高靶向miR-423-5p减轻ox-LDL诱导的HUVEC炎症反应和凋亡。
综上所述,ox-LDL引起HUVEC中LncRNA DANCR表达降低,过表达LncRNA DANCR通过靶向下调miR-423-5p可促进HUVEC存活,改善ox-LDL诱导的HUVEC炎症反应和凋亡,为基于HUVEC损伤的AS治疗提供了新的方向。