CXC亚家族趋化因子13在乳腺癌组织中表达及其调控网络的生物信息学分析
2021-09-18李英蕊李琴解军冯皓楠郭璇傅松涛
李英蕊,李琴,解军,冯皓楠,郭璇,傅松涛,4
1.山西医科大学基础医学院生物化学与分子生物学系,山西太原030000;
2.首都医科大学附属北京友谊医院肿瘤科,北京100021;
3.山西医科大学基础医学院病理学与病理生理学系,山西太原030000;
4.山西省生物医药健康研究生教育创新中心,山西太原030000
随着传统放化疗、内分泌治疗及靶向药物的应用,乳腺癌患者5年生存率得到极大提高,但乳腺癌发病率及死亡率仍在增长[1]。从分子水平研究乳腺癌机制,寻找新的治疗方法,是探索乳腺癌治疗的重要方向。肿瘤细胞的增殖、浸润由多种生长因子、趋化因子及细胞因子通过复杂的交互作用介导,其中趋化因子发挥重要的趋化作用,在多种炎症反应中激活特异性白细胞[2]。CXC亚家族趋化因子13(CXC subfamily chemokine 13,CXCL13)在肿瘤细胞生存及癌症转移中发挥重要的作用[3],CHU等[4]发现了两种对乳腺癌起关键作用的分子,CXCL13为其中之一。近来有研究表明,CXCL13可促进乳腺癌的进展及淋巴结转移[5-6]。
本文采用生物信息学方法分析CXCL13在乳腺癌组织中的表达及潜在的调控网络,为进一步探讨CXCL13在乳腺癌中的作用机制提供参考。
1 数据来源及分析
基于在线数据库分析乳腺癌组织中CXCL13。检索Oncomine数据库(https://www.oncomine.org/)分析CXCL13在乳腺癌组织中表达,获得CXCL13在多种乳腺癌组织中的转录水平数据,并对GLÜCK等[7]、CURTIS等[8]的研究结果及TCGA数据库中的乳腺癌组织转录组数据进行分析;检索c-BioPortal数据库(http://www.cbioportal.org/)分析乳腺癌样本中CXCL13基因的变化,对频率变化前50个CXCL13邻近基因用FunRich软件进行基因本体论(Gene Ontology,GO)的生物学过程(biological processes,BP)、细胞组分(cellular component,CC)及分子功能(molecular function,MF)分类分析,用京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)进行信号通路分析,进一步绘制并分析频率改变较高的前15个基因的生物互作网络;检索LinkedOmics数据库(http://www.linkedomics.org/login.php),使用皮尔森相关系数分析与CXCL13变化有关的基因,并对正相关的基因集进行GO及KEGG分析;使用基因集富集分析(gene set enrichment analysis,GESA)CXCL13正相关基因集在乳腺癌中的激酶、转录因子及miRNA的靶点网络。
2 结果
2.1 CXCL13在乳腺癌中的表达 结果显示,乳腺癌组织中CXCL13基因mRNA转录水平显著高于正常乳腺组织(P<0.01),虽差异倍数小于3,但CXCL13基因mRNA转录频率改变排在前6%;乳腺癌组织中CXCL13基因拷贝数变异总体高于正常乳腺组织,基因拷贝数变异排在前25%,但差异无统计学意义(P>0.05)。见图1。

图1 不同乳腺癌组织与正常组织中CXCL13基因mRNA转录水平和基因拷贝数的差异Fig.1 Difference of various breast cancer and normal tissues in CXCL13 gene mRNA transcription levels and gene copy numbers
2.2 CXCL13在乳腺癌中基因变化的频率及类型 结果显示,1 101例乳腺癌患者中23例发生CXCL13分子改变,占2.1%,其中17例为扩增突变,占1.5%,表明扩增是CXCL13在乳腺癌中最常见的突变类型,见图2。

图2 TCGA数据库中CXCL13基因变化情况Fig.2 Variation of CXCL13 gene in TCGA database
2.3 CXCL13变化在乳腺癌中的生物互作网络 经GO分类分析,CXCL13邻近50个发生突变的基因主要参与细胞通讯、信号转导等过程,定位于细胞质或细胞膜,涉及G蛋白偶联受体(G protein-coupled receptor,GPCR)活化、GTP酶活化、丝/苏氨酸蛋白酶激活等功能;经KEGG通路分析,这些基因富集在GPCR结合配体、肽受体结合配体及GPCR信号传导过程。见表1。频率变化较高的前15个邻近基因生物互作网络中,PTK2占25.6%,ERBB2占17.8%,NFAGT占16.3%,见图3。
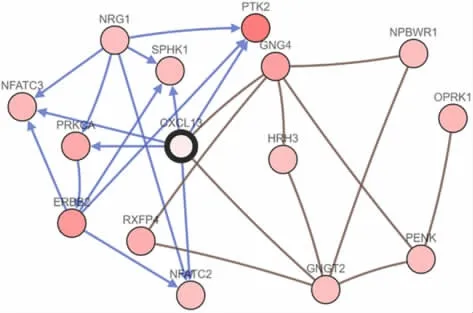
图3 乳腺癌中CXCL13与邻近基因生物相互作用网络Fig.3 Interaction network of CXCL13 gene and its adjacent genes in breast cancer

表1 乳腺癌中CXCL13邻近变化基因的富集分析Tab.1 Enrichment analysis of changed genes adjacent to CXCL13 gene in breast cancer
2.4 CXCL13在乳腺癌中的功能网络 富集分析显示,1 093例乳腺癌患者的测序数据中,5 579个基因与CXCL13变化呈正相关,3 591个基因呈负相关[错误发现率(false discovery rate,FDR)<0.01],见图4。经GO、KEGG分析,CXCL13正相关基因主要定位于细胞膜、免疫突触及受体复合体;主要参与适应性免疫反应、对趋化因子的反应、T细胞激活等过程,涉及细胞因子受体结合、活化等功能;主要富集在原发性免疫缺陷、T细胞受体信号通路、趋化因子信号通路、细胞因子与受体相关作用、造血干细胞谱系等过程。见图5。

图4 乳腺癌中与CXCL13相关的差异表达基因Fig.4 Differentially expressed genes associated with CXCL13 in breast cancer

图5 乳腺癌中CXCL13共表达基因的GO注释BP(A)、CC(B)、MF(C)及KEGG信号通路(D)分析(FDR<0.03)Fig.5 GO annotation of biological process(A),cell component(B),molecularfunction(C)and KEGG signaling pathway(D)of genes co-expressed with CXCL13 in breast cancer(FDR<0.03)
2.5 CXCL13在乳腺癌中的激酶、转录因子及miRNA的靶点网络CXCL13正相关基因集最重要的3个激酶靶点网络为FYN、LYN及LCK,均属于src家族酪氨酸激酶;miRNA靶点网络包括miR-146a和miR-519E等;转录因子靶点网络与NF-κB通路有关,包括V$NFKAPPAB_01和GGGNNTTTCC_V$NFKB_Q6_01。见表2。

表2 CXCL13正相关基因集的激酶、转录因子和miRNA的靶点网络Tab.2 Target networks of kinases,transcription factors and miRNAs of gene clusters positively related to CXCL13
3 讨论
趋化因子协调细胞迁移能力被恶性肿瘤不同的生物作用网络利用,但肿瘤的进展与肿瘤相关趋化因子表达的关系研究较少。PANSE等[9]发现,CXCL13在乳腺癌组织中过表达,转移性乳腺癌患者血清中该细胞因子表达升高,表明CXCL13在乳腺癌的进展中发挥作用,其可能作为一个有效的治疗靶点或恶性肿瘤的诊断指标。BISWAS等[10]研究发现,CXCL13刺激乳腺癌细胞可促进乳腺癌细胞的上皮间质转化,其可作为乳腺癌的不良预后指标。
本研究对TCGA数据库中1 101份临床样本的转录测序数据进行分析,证实CXCL13基因mRNA转录水平及拷贝数变异程度在癌组织中高于正常乳腺组织,CXCL13作为一种潜在的诊断及预后标志物值得进一步进行临床验证。基因拷贝数变异是基因组变异的主要来源,对癌症的发生发展有一定的影响。SAFONOV等[11]研究表明,拷贝数变异与肿瘤的免疫浸润呈正相关。本研究证实,CXCL13在乳腺癌中的拷贝数变异增加,其突变类型主要是扩增。因CXCL13具有重要的生理学功能,其基因改变可能造成多个下游信号通路的改变,涉及的相关功能网络包括GPCR的结合及信号传递,肽配体受体结合等过程,这与CXCL13的生理功能一致[12-13]。
本研究前期发现,神经调节蛋白1(neuroregulin 1,NRG1)激活可导致人类表皮生长因子受体-2(human epidermal growth factor receptor-2,HER2)阳性乳腺癌曲妥珠单抗原发耐药,NRG1可能作为预测曲妥珠单抗原发耐药的一个分子标记物[14]。本次分析也发现CXCL13与HER2及NRG1的变异有很大相关性,且生物互作图显示,这3个基因有很多相同作用靶点,提示CXCL13可能是导致曲妥珠单抗耐药的一个因素,值得进一步实验确证。有学者报道,CXCL13与CXC亚家族趋化因子受体-5(CXC chemokine receptor-5,CXCR5)相互作用遵循经典的GPCR激活模式[13],本文中CXCL13参与原发性免疫缺陷、T细胞受体信号通路、趋化因子信号通路、细胞因子与受体相关作用及造血干细胞谱系等过程,与前期报道一致。这些发现对探讨CXCL13在疾病中功能失调及癌症的发生机制具有重要意义。
基因组不稳定性及突变是癌细胞的基本特点,激酶及相关的信号通路可帮助稳定和修复基因组DNA[15-16]。本研究发现,在乳腺癌中CXCL13的激酶靶点网络中包括FYN激酶;XIE等[17]研究发现,FYN在乳腺癌中过表达,敲减FYN的表达可抑制乳腺癌细胞的增殖、迁移及浸润,并证明FYN可通过PI3K/AKT及ERK/MAPK通路诱导乳腺癌细胞的上皮间质转化。FYN在癌细胞对某些抗癌药物的耐药性或敏感性中具有重要意义;ELIAS等[18]研究发现,FYN在他莫昔芬耐药的乳腺癌细胞系中上调,并证明其在耐药机制中起关键作用。有研究表明,FYN参与T淋巴细胞的发育及激活[19]。SUGIE等[20]证明,FYN参与抗原特异性激活T细胞过程,抑制FYN严重损害T细胞反应。乳腺癌中CXCL13可能通过FYN激酶调控乳腺癌的发生发展或激活T细胞抗肿瘤反应,具体机制有待进一步验证。
转录因子对于细胞发育至关重要,转录因子包括多种蛋白质,它们可与顺式作用元件相互作用激活或抑制转录过程,恶性肿瘤的发生可能依赖于正常细胞发育程序,但叠加了潜在的基因变异。本研究发现,乳腺癌中CXCL13呈正相关变化的转录因子靶点网络中包含NF-κB通路。NF-κB途径在先天和适应性免疫、细胞增殖和存活、炎症和肿瘤发展中起关键作用[21-22]。常用的化疗及放疗方法激活NFκB,导致侵袭性乳腺癌的发展,表现出对化疗、放疗及内分泌治疗的耐药性[23-24]。CXCL13与受体结合可能增强NF-κB配体在乳腺癌细胞中的表达[10,25],它们之间的作用关系有待进一步探究。
本研究发现,CXCL13miRNA靶网络中包括miR-146a,其为一种免疫系统调节剂,与癌症、病毒感染及自身免疫性疾病等多种疾病相关[26]。CHENG等[27]发现,结直肠癌干细胞可通过分泌负载miR-146a的致癌外泌体,编码结直肠癌细胞的干细胞特性及致瘤性。SUBRAMANIAN等[28]分析乳腺癌患者与健康成年人血浆中miR-146a表达,发现乳腺癌患者血浆中miR-146a水平明显升高,可能对乳腺癌具有诊断作用。TAN等[29]认为,miR-146a是多药耐药性肺癌的潜在治疗靶标。ASSUNTA等[30]研究表明,NF-κB与miR-146a协同作用使病毒DNA在细胞中的积累和传染颗粒产生。NF-κB与miR-146a在乳腺癌的发生发展中均有变化,两者之间的相互作用也是下一步研究的重点。
本研究基于TCGA数据库,采用生物信息学在线工具,对肿瘤数据进行目标基因分析,使得大规模的乳腺癌基因组研究和随后的功能研究成为可能。但本研究仍存在局限性,如乳腺癌测序数据来源分组不同,患者的遗传背景及病因可能存在显著差异,转录组测序仅能检测到静态突变,不能直接提供蛋白质活性或表达水平的信息。因此,需进一步使用临床样本进行相关的分子生物学研究。
综上所述,CXCL13在乳腺癌组织中高表达,该变化对GPCR的激活、信号传递及T细胞激活等多个步骤具有深远的影响;CXCL13与几种肿瘤相关激酶(如FYN)、转录因子(如NF-κB)及miRNA(如miRNA-146a)特异相关。本研究为CXCL13在乳腺癌发生中的重要作用及其作为乳腺癌标志物的潜能提供了参考。
