胶孢炭疽菌细胞骨架荧光标记菌株的构建
2021-09-14刘娜刘世科王倩男
刘娜 刘世科 王倩男
(海南大学热带作物学院 海南省热带生物资源可持续利用重点实验室,海口 570228)
胶孢炭疽菌(Colletotrichum gloeosporioides)寄主广泛,其多个不同的生理小种能够侵染诸如橡胶树、香蕉、火龙果、香瓜、西瓜等多种经济作物,是造成农业减产的主要病害之一。深入研究胶孢炭疽菌的致病力机制,有助于新型防控措施的开发。
细胞极性的确立和维持在真核生物的形态发生过程中起着重要的作用。丝状真菌的分生孢子萌发后,菌丝呈现出明显的极性生长现象[1-3]。在这个极性生长过程中,细胞壁及细胞膜前体物质、细胞器以及囊泡等不断运输汇集到菌丝顶端的特殊结构(spitzenkörper,Spk)[4]。Spk由密集的囊泡、细胞骨架、核糖体及其他多种物质组成[5-6];与此同时,Spk介导的菌丝顶端胞外物质分泌也与真菌的生长发育及致病力密切相关[7-8]。虽然目前关于Spk的形成及其机制还尚不清楚,但是普遍认为细胞骨架在其中起着重要作用。细胞骨架广泛存在于真核生物的细胞质及细胞核中。细胞骨架在细胞分裂、细胞生长等多项生理活动中具有重要的功能。细胞质骨架主要包括微丝、微管和中间纤维,其中关于微丝和微管的研究较多。微丝(microfilament)是由单体肌动蛋白(actin)呈螺旋状排列组装而成;微管(microtubule)是由α-微管蛋白(tubulin)和β-微管蛋白通过非共价键连接组成的中空管状结构。在细胞内,微丝和微管均处于聚合和解聚的高度动态循环中,而这一动态过程分别受一系列微丝结合蛋白(actin binding protein,ABP)和微管结合蛋白(microtubule associated protein,MAP)的精密调 控[9-10]。至今为止,尚未有关胶孢炭疽菌细胞骨架研究的相关报道。
Lifeact是来自出芽酵母(Saccharomyces cerevisiae)ABP140(actin binding protein 140)一段17个氨基酸的多肽,其序列被广泛用作真核生物细胞内微丝骨架标记探针[11-12],已有研究结果表明Lifeact能很好的标记出稻瘟菌、灰霉菌等真菌细胞中的微丝骨架结构[13-14]。而在微管研究中发现,鼠微管相关蛋白MAP4(microtubule associated protein 4)的微管结合结构域MBD(microtubule binding domain)能很好的标记出植物细胞内微管的结构[12,15]。因此,本研究构建了增强型绿色荧光蛋白EGFP(enhanced green fluorescent protein)与Lifeact及MBD的融合表达载体,将其转化胶孢炭疽菌并获得了相应的重组菌株。与此同时,还在胶孢炭疽菌β-微管蛋白(β-tubulin)编码基因(CgTUB1)后面原位敲入了EGFP的编码序列,构建了CgTUB1-EGFP标记菌株。显微观察及生理表型分析结果表明,所构建的Lifeact-EGFP及CgTUB1-EGFP的重组菌株的微丝及微管骨架标记清晰,为进一步研究胶孢炭疽菌细胞骨架的动态变化及骨架结合蛋白的功能奠定基础。
1 材料与方法
1.1 材料
1.1.1 供试菌株 橡胶树胶孢炭疽菌野生型菌株由本实验室分离保存,并在前期工作中完成该菌株的基因组测序工作。
1.1.2 实验试剂 实验所用基因表达载体pPgTH由本实验室改造,以pMD19-T为骨架,在Pst I和Xba I位点处连入构巢曲霉3-磷酸甘油醛脱氢酶基因的启动子PgpdA,在Sac I和EcoR I处连入构巢曲霉色氨酸合成基因trpC的终止子(TtrpC)及潮霉素磷酸转移酶抗性基因(HPT)。PCR引物合成及DNA测序由华大基因及擎科生物公司完成。DNA聚合酶、DNA Marker及大肠杆菌感受态均购自北京全式金公司,限制性内切酶及T4 DNA连接酶购自Thermo Scientific公司,胶回收试剂盒、质粒提取试剂盒购自天根公司。羧苄青霉素、卡那霉素购自索莱宝公司,潮霉素购自Roche公司,其它化学试剂采用国产分析纯。胶孢炭疽菌的培养使用土豆浸膏培养基 (PDA,PDB)及基本培养基(minimal medium)。
1.2 方法
1.2.1 微丝骨架标记载体的构建 采用随机重组原理敲入标记基因,具体策略如图1-A所示,将Lifeact与绿色荧光蛋白EGFP的融合编码序列Lifeact-EGFP连入基因表达载体pPgTH的Xba I和Sac I位点,构建微丝标记载体。
1.2.2 微管骨架标记载体的构建 分别采用随机重组原理及同源重组原理等两种策略构建两种微管骨架标记载体[16]。(1)将鼠MAP4的微管结合结构域MBD与绿色荧光蛋白EGFP的融合编码序列MBDEGFP连入载体pPgTH的Xba I和SacI位点,构建微管标记载体(图1-B)。(2)将胶孢炭疽菌β1 tubulin DNA序列CgTUB1的3'端序列(去除终止密码子,853 bp)连入载体pPgTH的Xba I和Kpn I位点,将EGFP编码序列连入Kpn I和Sac I位点,再使用引物CgTub-SF/hph-splR将这两个片段与潮霉素抗性基因编码序列(HPT)的5'端序列PCR扩增备 用,所得产物为左侧敲除片段;之后将HPT的3'端序列与CgTUB1的下游序列(flanking region,804 bp)进行融合PCR扩增备用,所得产物为右侧敲除片段;最后将两个敲除片段共同转化野生型菌株,将CgTUB1 DNA的3'端序列作为上游同源臂、CgTUB1的下游序列作为下游同源臂,利用同源重组原理将EGFP编码序列原位敲入CgTUB1的下游,构建敲入重组菌株(图1-C)。

图1 细胞骨架荧光标记载体原理图Fig. 1 Diagram of fluorescent labeled cytoskeleton vectors
1.2.3 原生质体转化 橡胶树胶孢炭疽菌原生质体的制备及转化参照文献[3]所述方法进行。在转化前,将Lifeact-EGFP、MBD-EGFP及CgTUB1-EGFP标记载体用Pst I进行酶切线性化处理。
1.2.4 重组菌株的鉴定 本实验中采用潮霉素(300 μg/mL)筛选转化子。待转化、再生培养后,将阳性转化子转接到PDA培养基进行扩大培养,并提取其基因组DNA进行PCR鉴定。对于转化过Lifeact-EGFP或MBD-EGFP的转化子,以其基因组为模板,分别扩增其基因组中是否已经转入目标序列。对于CgTUB1-EGFP转化子,为了检测敲入重组菌株中EGFP编码序列是否重组到CgTUB1基因后面,本实验中采用两轮PCR检测方法鉴定转化子,在重组片段中的上游同源臂的外侧及EGFP序列中设计一对检测引物CgTub-JCF/EGFP-R;同理,再在下游同源臂的外侧及抗性基因HPT序列中设计一对检测引物HPT-JCF/CgTub-JC3R;通过PCR扩增检测重组片段是否正确整合到目标基因的位点(表1)。在获得以上所述3种阳性重组菌株后,通过单孢分离对筛选到的重组菌株进行分离纯化。
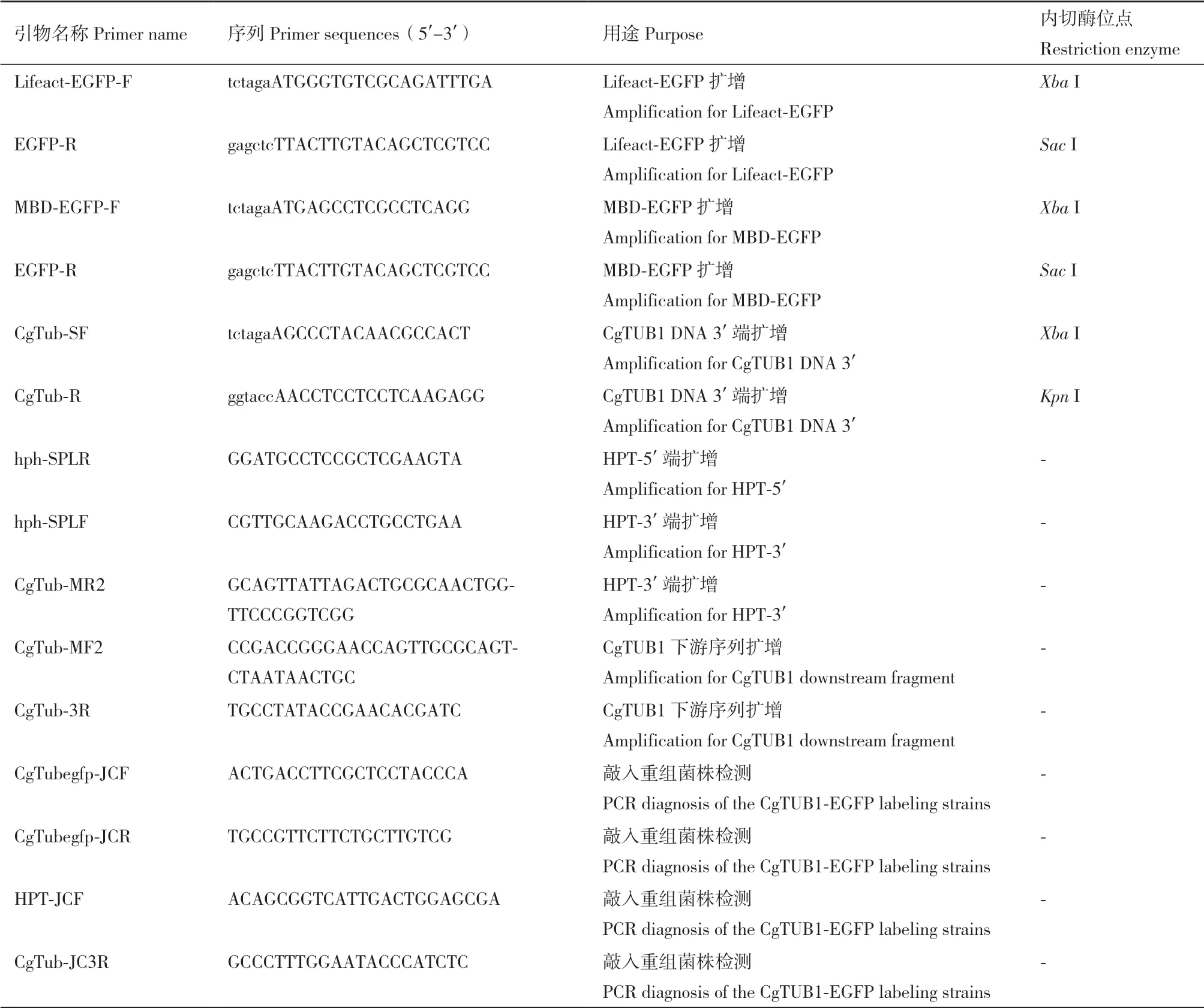
表1 本实验所用引物Table 1 Primers used in this study
1.2.5 激光扫描共聚焦显微镜观察重组菌株中细胞骨架 将获得的纯合体重组菌株接种于PDB培养基中,28℃震荡培养2 d后,过滤收集孢子,用ddH2O洗涤2遍后重悬,再于激光共聚焦显微镜下观察孢子内微丝或微管骨架的分布情况。与此同时,将上述孢子接种于新鲜的PDB培养基中,震荡培养24 h后,于激光共聚焦显微镜下观察菌丝内微丝或微管骨架的分布情况。所有荧光图片均采自Leica SP8激光扫描共聚焦荧光显微镜。EGFP由Argon 激光器488 nm通道激发,检测的发射波长范围是505-540nm。静态层扫图片的步距为0.5 μm。所有图片均通过Image J 软件进行后期叠加及处理。
1.2.6 生长速率及产孢量检测 参照文献[3]所述方法进行。
1.2.7 致病力分析 参照文献[3]所述方法进行。每个处理共包含30片叶片,分为3组。本实验共重复2次。
2 结果
2.1 重组菌株的鉴定
胶孢炭疽菌原生质体转化后分别获得了多个具有潮霉素抗性的阳性转化子。接着将转化子转接到PDA平板进行培养,提取其DNA,并利用PCR检测各转化子。实验中共获得了多个Lifeact-EGFP及MBD-EGFP表达载体转化子;分别随机挑取4个转化子,并用PCR扩增检测其基因组中是否已转入Lifeact-EGFP或MBD-EGFP片段。结果这两组转化子均能检测到目标片段(图2-A,B),表明标记重组菌株构建成功,分别将这两组重组菌株命名为PgpdA∷Lifeact-EGFP和PgpdA∷MBD-EGFP。与此同时,挑取了10个CgTUB1-EGFP基因敲入重组转化子,并对转化子进行两轮PCR检测。结果有3个转化子能够同时成功扩增出1 722 bp、1 648 bp大小的目标条带(图2-C,D),且PCR产物测序结果也与预期结果一致,表明这3个转化子均已在CgTUB1的DNA序列后面敲入了EGFP编码序列,因此将这3个转化子命名为CgTUB1-EGFP。之后通过单孢分离,分别获得了PgpdA∷Lifeact-EGFP、PgpdA∷MBD-EGFP和CgTUB1-EGFP的纯合体重组菌株。

图2 细胞骨架荧光标记重组菌株的PCR鉴定Fig. 2 PCR diagnosis of cytoskeleton labeling recombinant strains
2.2 重组菌株细胞骨架观察
将上述获得的PgpdA∷Lifeact-EGFP纯合体重组菌株制片后置于共聚焦显微镜下观察其微丝骨架。结果发现在其孢子及菌丝细胞内均能检测到明显的荧光,并且与只表达EGFP的对照菌株相比,重组菌株孢子内微丝主要以点状信号分布,也有少量丝状结构存在(图3)。在菌丝中,可在其顶端观察到明显的微丝聚集现象,呈明显的极性分部;此外在细胞中间部位,也能观察到丝状结构(图4)。以上结果表明Lifeact-EGFP能够较好的标记胶孢炭疽菌的微丝骨架。随后,选择了其中一株荧光强度较高的重组菌株进行后续研究。
将上述获得的PgpdA∷MBD-EGFP及CgTUB1-EGFP纯合体重组菌株制片后置于共聚焦显微镜下观察其微管骨架。结果发现PgpdA∷MBD-EGFP菌株细胞内荧光强度极低,难以观测到其孢子及菌丝内部的微管骨架结构(结果未展示);而CgTUB1-EGFP标记重组菌株的孢子及菌丝细胞内均能检测到明显的荧光。如图3所示,与对照菌株相比,标记菌株孢子内的微管主要呈点状和丝状结构。在菌丝内部微管也呈现出较明显的极性分布现象,此外,微管主要以点状和丝状分布于菌丝中间(图4)。以上结果表明CgTUB1-EGFP能较好的标记微管骨架。随后,选择了其中一株荧光强度较高的CgTUB1-EGFP重组菌株进行后续研究。

图3 荧光标记重组菌株孢子中的细胞骨架结构Fig. 3 Cytoskeleton structure in conidiophores of fluorescent labeled recombinant strains

图4 荧光标记重组菌株菌丝中的细胞骨架结构Fig. 4 Cytoskeleton structure in the hypha of fluorescent labeled recombinant strains
2.3 重组菌株生长速率及产孢量检测
在PDA培养基中培养时,PgpdA∷Lifeact-EGFP、CgTUB1-EGFP重组菌株与野生型菌株相比,其菌落生长速率及菌落形态都没有显著差别(图5-A),在培养4 d后,菌落直径均约为6 cm。与此同时,在液体PDB培养基中培养3 d后,重组菌株的产孢量与野生型相比也没有显著变化,孢子产量都约为35×105个/mL(图5-B)。
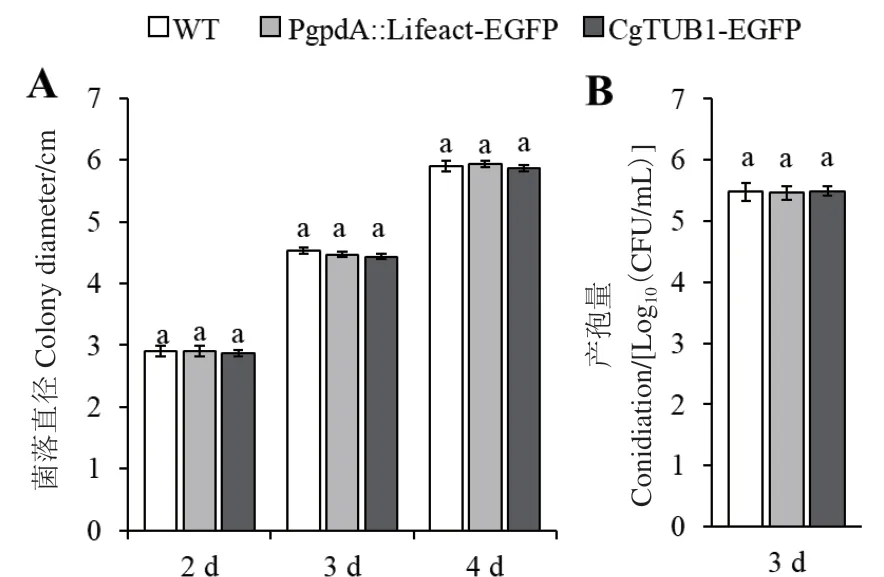
图5 野生型菌株与重组菌株菌落直径及产孢量Fig. 5 Clony diameters and conidiations of WT and mutant strains
2.4 重组菌株致病力分析
将PgpdA∷Lifeact-EGFP、CgTUB1-EGFP重组菌 株与野生型菌株同时接种橡胶树叶片。在接种后,重组菌株与野生型菌株均能侵染橡胶树叶片并造成病斑;与此同时,在接种2 d及3 d后,重组菌株与野生型菌株形成的病斑直径均约为10 mm及20 mm(图6)。表明重组菌株与野生型菌株致病力没有明显差异。
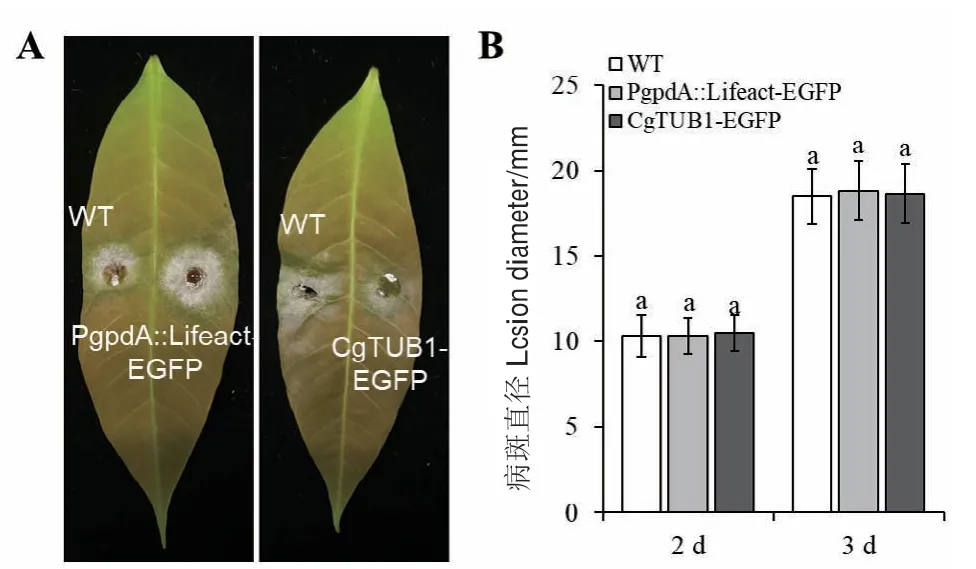
图6 接种野生型菌株与重组菌株后橡胶树叶片的发病情况及病斑直径Fig. 6 Virulence assay on rubber tree leaves and lesion diameter after inoculation with WT and recombinant strains
3 讨论
细胞骨架广泛存在于真核生物细胞中,并在其形态建成、物质运输与极性生长等方面有着重要的功能。丝状真菌的生长和发育主要依赖菌丝尖端的极性生长,而这种极性生长受到细胞内骨架的严格调控[17-18]。近年来,越来越多的研究揭示了细胞骨架的结构及其动态变化在丝状真菌形态发生与致病过程中的作用[19-21]。因此,要研究胶孢炭疽菌中细胞骨架结构及其动态变化及相关蛋白功能,对胞内细胞骨架进行荧光标记和活体细胞观察是必要的。由于actin-GFP(green fluorescent protein)难以加入到聚合的微丝中,若直接对actin进行荧光标记还会导致单体肌动蛋白被标记且呈现较强荧光背景[22]。此外,actin-GFP的表达严重影响了盘基网柄菌细胞(Dictyostelium)的胞质分裂等生理生化活动[22]。目前对于微丝骨架的活体细胞成像来说,主要通过微丝结合蛋白来标记F-actin,如拟南芥AtFIM1的微丝结合结构域ABD2[23],以及酵母ABP140的一段仅17个氨基酸的保守结构域Lifeact[24-29]。Delgado-Alvarez的相关研究表明,通过对粗糙脉孢霉(Neurospora crassa)多个微丝结合蛋白及其结构域与绿色荧光蛋白GFP进行融合表达,构建了多种荧光标记菌株:Fimbrin-GFP,ARP3-GFP,tropomyosin-GFP,Lifeact-GFP;结果发现不同的荧光标记菌株能标记出菌丝内不同的微丝骨架结构,但Lifeact-GFP能较好的标记出菌丝不同部位的多种结构[30]。此外,Lifeact也被广泛用于其他真菌胞内微丝骨架的标记[13,31-33]。在此基础上,本研究构建了PgpdA∷lifeact-EGFP表达载体,通过胶孢炭疽菌原生质体转化成功构建了重组菌株,并对微丝骨架的三维立体结构进行了活体观察。结果表明,在胶孢炭疽菌中微丝主要以点状信号分布于孢子中,以点状和丝状结构分布于菌丝中,且呈现出明显的极性分布现象,这一结果也与其它真菌中观察到的结果相类似[13,30]。在丝状真菌中,点状微丝结构是细胞内吞作用的标志[34]。稻瘟菌中菌丝顶端点状微丝结构的减少会导致胞吞作用明显延迟,效应因子分泌受阻,致病力受到严重的影响[35]。
而对于微管骨架,常用小鼠MAP4的MBD与荧光蛋白融合进行标记[15,36],或直接用tubulin与荧光蛋白融合来标记动植物细胞中的微管骨架[37];但是在不同生物的不同组织中,两种标记方法的效果不尽相同。在构巢曲霉(Aspergillus nidulans)中,利用GFP-TUA(α-tubulin)菌株来观察菌丝中的微管骨架结构[38-39]。在粗糙脉孢霉中,β-tubulin-GFP被用来标记胞内的微管骨架[40]。在禾谷镰刀菌(Fusarium graminearum)中,多用FgTub1-GFP及FgTub2-GFP来标记胞内的微管骨架[21,41-42]。在本研究中分别使用了两种方法对胶孢炭疽菌的微管结构进行标记:构建了PgpdA∷MBD-EGFP-TtrpC表达载体与CgTUB1基因位点后EGFP敲入载体,并通过原生质体转化成功获得重组菌株。结果表明,MBD-EGFP标记重组菌株中荧光强度较低,而CgTUB1-EGFP标记重组菌株荧光强度较高;与此同时,在CgTUB1-EGFP重组菌株孢子和菌丝中都可观察到明显的点状和丝状信号,且在菌丝尖端的微管骨架也呈现出一定的极性分布现象。
为了检测两种骨架标记重组菌株是否会对胶孢炭疽菌正常的生理活动产生影响,本研究分别选择了两株荧光强度较高的PgpdA∷lifeact-EGFP和CgTUB1-EGFP重组菌株,对其生理表型进行了分析。结果发现这两种细胞骨架的荧光标记并不影响胶孢炭疽菌的生长、产孢及致病力,表明本研究所构建的重组菌株能够用于后续分析。
丝状真菌细胞顶端极性的建立、维持都需要细胞骨架的参与,其结构的动态变化在细胞生长、胞内物质运输以及外泌过程中扮演着至关重要的角色,因而在其致病过程中也发挥着重要的作用[43]。细胞骨架结构由肌动蛋白、微管蛋白及很多相关蛋白(ABPs,MAPs)共同调控。且微管蛋白还是很多真菌杀菌剂的作用靶点[42]。鉴于此,对于胶孢炭疽菌中微丝微管骨架的荧光标记将有利于推动对其菌丝生长及致病过程中细胞内骨架分布、动态变化及其相关蛋白功能,以及对杀菌剂作用机理的研究。
4 结论
本研究构建了胶孢炭疽菌微丝及微管骨架的标记重组菌株,为今后深入研究胶孢炭疽菌微丝和微管骨架的结构、动态变化、以及细胞骨架相关蛋白的功能奠定了良好的基础。
