苜蓿抗旱性分子研究进展
2021-09-14李倩江文波王玉祥张博庞永珍
李倩 江文波 王玉祥 张博 庞永珍
(1. 中国农业科学院北京畜牧兽医研究所,北京 100193;2. 新疆农业大学草业与环境科学学院,乌鲁木齐 833400)
苜蓿属(Medicago)植物属豆科蝶形花亚科,包括83个种,目前研究较为广泛的饲草主要包括模式植物蒺藜苜蓿(Medicago truncatula)、黄花苜蓿(Medicago falcata L.)和紫花苜蓿(Medicago sativa L.)[1]。其中紫花苜蓿是世界上种植面积最为广泛且最具经济价值的豆科栽培牧草[2],全世界的紫花苜蓿种植面积约 3.22×107hm2,而我国苜蓿种植面积约为3.77×106hm2[3],主要分布在西北、华北、东北各省区,南方各地也开始栽种[4]。黄花苜蓿在我国分布广泛,大部分为野生资源,具有较好的抗性,适口性好,为牛、羊提供丰富的营养,可与紫花苜蓿杂交形成抗性强、利用价值更高的杂花苜蓿,是苜蓿抗性育种的重要基因库[5]。杂花苜蓿抗逆性强,杂种优势显著,表型多样性丰富,具有良好的选育潜质,可为进一步选育不同优良特性的新品种提供适宜的育种材料[6];蒺藜苜蓿是一年生植物,其基因组小且自花授粉,是研究豆科牧草的模式植物[7]。尽管苜蓿品种能够适应水分有限的地区,但水分亏缺仍然影响紫花苜蓿的产量。因此,通过提高苜蓿抗旱能力进而提高草产量已成为紫花苜蓿研究的重要方向[8-9]。
干旱胁迫严重影响苜蓿的生长、发育及产量。据估计,干旱和半干旱地区约占全球总面积的30%[10]。如今,人们已充分认识到干旱胁迫严重制约了苜蓿产业的发展,并通过分子生物学手段和技术对紫花苜蓿加以改良。因此,以常规育种为基础,借助于生物技术改良抗旱性状,培育高产、高抗、优质的新品种,是一种更直接、有效的方式。本文将从分子水平上对4种苜蓿的抗旱性进行分析,详细总结目前苜蓿抗旱基因及抗旱分子育种方面相关的研究进展,以期为苜蓿的抗旱品种改良提供参考。
1 分子学水平的研究进展
1.1 组学研究进展
随着对苜蓿的研究不断深入,苜蓿的抗性育种和品种改良已成为当下研究的重点。但由于黄花苜蓿的基因组信息尚未公布,只能参考豆科模式植物蒺藜苜蓿和紫花苜蓿的基因组信息辅助研究黄花苜蓿的抗旱性。在紫花苜蓿(新疆大叶)的基因组已经公布的情况下[11],结合转录组测序技术,对苜蓿的抗逆基因挖掘和生物信息学分析具有重要的参考价值。
在紫花苜蓿基因组公布之前,有多个关于紫花苜蓿转录组测序的研究报道,为研究紫花苜蓿抗旱分子机理提供参考。通过转录组测序技术揭示了苜蓿内源NO胁迫耐受的相关基因和途径[12],Luo等[13]利用三代测序技术PacBio构建了第一个紫花苜蓿根尖在连续甘露醇处理下的全长转录组数据库,共鉴定了8 016个受甘露醇调节的差异表达基因。还通过代谢分析发现渗透相关基因,筛选出来的基因可作为未来研究抗旱的关键候选基因。研究人员于2020年利用三代PacBio、Bionano及HIC技术,结合最先进组装策略破译了紫花苜蓿栽培种“中苜1号”基因组,并对世界范围内的紫花苜蓿核心种质进行重测序并获得许多重要农艺性状关联候选区域[14]。另一项研究通过GWAS与基因分型的测序鉴定了紫花苜蓿与抗旱系数和叶片相对含水量等抗旱性状相关的位点[15]。Ray等[16]利用重测序技术检测了在干旱胁迫下紫花苜蓿生物量相关的数量性状位点,Wang[17]等也证明GWAS结合GBS是分析紫花苜蓿农艺性状的一种有效方法,其结果对进一步鉴定表型相关候选基因具有重要的参考价值。
为了筛选黄花苜蓿抗旱相关基因,研究人员通过构建抑制性差减杂交文库和转录组测序技术,挖掘了植物激素代谢相关信号途径中的关键基因,筛选了黄花苜蓿中干旱响应的特异基因[18]和一个受脱水干旱胁迫诱导显著上调表达的NAC转录因子MfNACsa[19],这些基因都可用于牧草和作物育种的候选基因。对干旱胁迫下黄花苜蓿转录组的sRNA测序,结果表明黄花苜蓿一方面通过调控膜蛋白来增加水分和离子的流动以调节渗透势,另一方面通过促进磷脂代谢,提高膜系统的稳定性,从而调控干旱[20]。
随着苜蓿基因组的公布以及测序技术的不断更新,多组学相结合分析苜蓿抗旱机理成为目前的研究趋势。通过转录组挖掘干旱逆境胁迫相关基因[21],发现了蒺藜苜蓿 R2R3-MYB 转录因子主要参与调控氧化还原反应、细胞信号转导以及转录调控等过程,从而抵御干旱等非生物胁迫[22]。Kang等[23]利用蒺藜苜蓿全基因组关联分析发现,醛脱氢酶家族蛋白与气孔密度具有显著相关性,说明该基因在干旱胁迫下严重影响气孔密度。为探究 miRNAs 在植物响应干旱胁迫中的作用,Wang等[24]通过Illumina高通量测序在蒺藜苜蓿中鉴定了miRNA家族的新成员发现,这些miRNA均可响应干旱胁迫。
1.2 功能基因研究
对苜蓿抗旱功能基因的研究主要集中在转录因子和基因表达调控方面,通过克隆抗旱相关基因,可阐明紫花苜蓿抗旱调控中的分子机制,并有望应用于分子改良中[25]。目前,一部分转录因子也已经被发现参与了苜蓿应答干旱胁迫(表1)。

表1 苜蓿来源的抗旱相关基因及其功能研究Table 1 Research on drought resistance related genes and their functions from Medicago
NAC转录因子是植物特有的一类转录因子,在植物生长发育、响应逆境胁迫途径中起重要作用。目前,紫花苜蓿中MsNAC2[26]、黄花苜蓿中MfNAC35[27]和MfNACsa[19]等基因都被成功克隆,基因表达模式均已明确,实验证明这些基因可能参与调控抗旱逆境胁迫的响应。MYB型转录因子参与植物对多种胁迫因子的反应。然而,在苜蓿中,只有少数几个MYB基因与胁迫反应相关。将紫花苜蓿的MsMYB2L基因在拟南芥中过表达时,干旱胁迫处理后发现,该基因的过量表达促进了脯氨酸、可溶性糖等渗透调节物质的合成,降低了脂质过氧化,从而提高了转基因拟南芥的抗旱性[28]。ERF转录因子是植物特有的一类转录因子,广泛参与植物多种逆境胁迫。苜蓿中MfERF049[29]、MfERF026、MfERF086[30]和MsERF11[31]被成功克隆,不同的非生物胁迫都可诱导这些基因的表达,表明这些基因可能参与了多种逆境胁迫的调控过程。Li等[32]克隆了MsZIP基因,过量表达该基因的烟草对渗透压胁迫的耐受性增强。王菊萍等[33]从蒺藜苜蓿幼苗中克隆了MtbHLH148 基因,发现该基因受自然脱水胁迫处理后表达量上调。目前,对于苜蓿中抗旱相关转录因子的研究,也只是将候选基因进行克隆奠定,并且证明这些基因受不同非生物胁迫的诱导,部分基因被转化进模式植物烟草或者拟南芥中进行功能验证,但均未转入本体植物苜蓿中进行功能验证。
除了转录因子,苜蓿来源的其它类型的基因也参与干旱等多种逆境胁迫反应(表2)。研究表明,紫花苜蓿来源的GDP-甘露糖3,5-表异构酶MsGME基因在拟南芥中异源表达时,可通过降低MDA含量、较少膜损伤,以及增加抗坏血酸的积累来增强其抗旱能力[34]。紫花苜蓿来源的玉米黄质环氧化 酶(MsZEP)[35]和Orange(MsOr)基 因[36]在烟草中表达时,可通过分别提高相对含水量、ABA 含量、类胡萝卜素含量,调节气孔导度及气孔开度,从而增强抗旱性。在烟草中过量表达黄花苜蓿的MfHyPRP[37]基因时,增加了肌醇、甜菜碱和棉籽糖的合成量,在干旱胁迫处理下,转基因烟草相对于未转基因对照具有更高的光合效率和更低的离子渗漏,对干旱的耐受性增强。在蒺藜苜蓿中对MtALDH7A1[38]和MtLEA5B[39]基因的克隆和深入研究证明这些基因对干旱生境的抗逆响应具有重要作用。另外的一项研究表明,在拟南芥中过表达苜蓿来源的低温驯化特异蛋白基因MtCAS31时,显著降低了转基因拟南芥的气孔密度,大大提高了转基因拟南芥的耐旱性[40]。
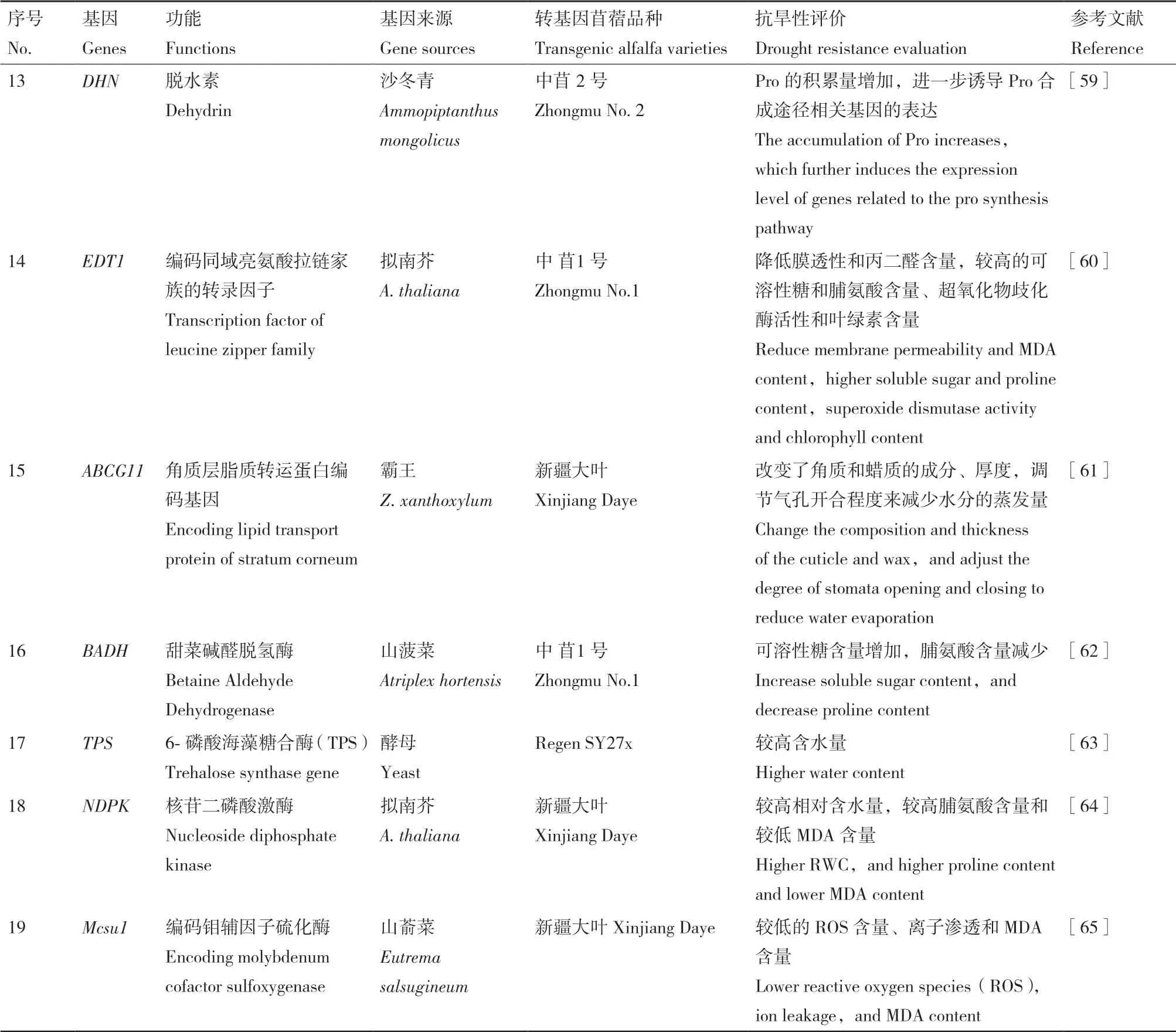
表2 续表 Continued

表2 用于提高紫花苜蓿抗旱性的基因及其作用Table 2 Genes used to improve drought resistance of alfalfa and their functions
以上研究大多通过同源克隆的方式研究苜蓿的关键转录因子或酶基因,并进行序列分析,阐明基因在不同组织中的表达模式;相对于对模式植物抗旱机制的研究而言,目前对苜蓿抗旱相关基因的功能验证和应用较为粗浅,大多数研究均是将相关基因通过生物技术转入其它模式植物或者紫花苜蓿/蒺藜苜蓿中,进行初步的功能分析,更具体的抗旱调控分子机理尚不清楚,还需要进一步的研究和探索。
2 利用基因工程进行分子育种
目前我国苜蓿育种工作中可供利用的优异种质资源严重缺乏,迫切需要发掘和创造新种质、新材料、新基因[41]。目前只有抗农达除草剂和低木质素含量的苜蓿新品种已经投放市场[42-43],但在我国审定登记的苜蓿品种中,仍无单纯运用生物技术培育成的紫花苜蓿品种。为了加快育种进程,克服传统育种缺陷,全世界育种学家们热衷于基因工程技术辅助的育种方法[44]。为了减轻苜蓿生产过程中的产量和品质损失,大力发展适应各种非生物胁迫的苜蓿新品种。自1986年苜蓿转基因技术首次取得成功后[45],该技术为苜蓿改良提供了一种有效的技术手段,目前很多抗旱相关基因均用于苜蓿抗旱的分子改良中,这些基因主要分为两类:转录因子和其它类型的基因。
将外源基因转入紫花苜蓿中,转基因植株通过减少膜损伤和渗透胁迫来提高耐旱性,这主要归因于增强叶片表皮蜡质、增厚角质层、减少水分流失和增强光合作用,进而提高脯氨酸含量、可溶性糖的含量、叶绿素含量、相对含水量和酶活性、降低丙二醛含量、蒸腾速率和活性氧含量。尽管不同基因在抗旱中分子机制可能有所不同,但它们都能通过渗透调节、离子调节和改变酶活性,从而提高转基因苜蓿的抗旱性。
3 展望
植物抗旱是一个非常复杂的机制,是受多基因控制的数量性状,是耐旱相关代谢途径以及其它多种代谢途径协同作用的结果[66],苜蓿的耐旱分子机理仍不清楚。目前有关苜蓿抗旱性的分子研究进展,主要集中在通过组学研究筛选抗旱相关基因,大多数研究均是将相关基因通过生物技术转入其它模式植物或者紫花苜蓿/蒺藜苜蓿中,进行初步的功能分析,更具体的抗旱调控分子机理尚不清楚,还需要进一步的研究和探索。关于苜蓿分子育种研究较为迟缓,也无单纯运用生物技术培育成的紫花苜蓿品种。目前热门的基因编辑技术可能对紫花苜蓿进行定向遗传改良,需要全面研究并解析紫花苜蓿耐旱相关代谢途径、耐旱相关基因及其表达模式。而通过传统育种也未培育出抗旱性强的紫花苜蓿材料和品种,所以通过分子生物学技术手段提高紫花苜蓿的抗旱性是进行紫花苜蓿分子育种的有效手段,但仍然存在一些问题。首先,缺乏系统完整的抗旱性鉴定评价标准;其次,转基因株系一般是基于人工模拟的环境-实验室或温室中进行的干旱胁迫,大多缺少相应的品种区域特性试验,很少投入到实际生产过程中。而区域试验是评价新品种适应性的最直接可靠的办法,需要设立和增加转基因紫花苜蓿区域试验站(点)的数量,从而使转基因的新品系得到大田验证并得以推广示范用于生产应用[67]。因此,进行紫花苜蓿育种是一项长期的工作。
苜蓿转基因的安全性也是一个备受关注的热点话题。首先,紫花苜蓿作为重要的豆科饲草主要应用于牲畜中,而不是直接应用于人类自身,相对而言也许较为安全。其次,相对于牧草品质性状的改良而言,将抗旱等抗逆性状基因转入紫花苜蓿中并不改变紫花苜蓿的品质性状,对动物和人类的食品安全的影响可能更小。最后,黄花苜蓿是目前很多紫花苜蓿育成品种的亲本材料,黄花苜蓿本身的抗旱能力很强,如果将黄花苜蓿的抗旱基因转入紫花苜蓿,在保证产量的前提下提高抗性,也许远比转入其它植物来源的外源基因更加安全可靠,这可能是进行紫花苜蓿抗旱性改良的最有效的策略,也是当前苜蓿抗旱育种的研究目标。
