金耳栽培基质物质降解与转化规律研究
2021-09-13曹瑶,李荣春,杨林雷,李梦杰,罗祥英,沈真辉,陆青青
曹瑶,李荣春,杨林雷,李梦杰,罗祥英,沈真辉,陆青青
摘要:【目的】探索金耳不同生長发育时间栽培基质内物质降解与转化的变化规律,了解金耳对碳源的降解利用特点,为栽培培养基配方设计提供理论基础。【方法】测定金耳8个生长时间点[未接种(0 d),以及接种后20、25、30、35、40、45和50 d]栽培基质中含水量、pH、培养料湿重及干重、子实体鲜重及干重、木质纤维素组分的降解量,分析不同生长阶段内各指标的变化情况。【结果】金耳生长发育周期内,含水量由60.27%下降至39.00%。接种后25 d pH由5.50降至4.55,之后pH基本稳定在4.20左右,总体为酸性条件。培养料干重由最初的735.00 g/袋降至530.00 g/袋,子实体鲜重产量为644.71 g/袋。总培养基失重率27.89%,其中,绝对生物学效率12.33%,呼吸作用消耗率15.56%。30~35 d为子实体生长第1次高峰期,40~45 d为子实体生长第2次高峰期。0~40 d呼吸作用随着金耳子实体生长不断增加。木质纤维素组分的吸收利用顺序依次为纤维素、木质素和半纤维素,其利用率分别为33.90%、32.79%和30.76%。生长前期(0~30 d)主要以非木质纤维素类物质为碳源,生长后期(30~50 d)主要以木质纤维素类物质为碳源;整个生长周期内所需的44.72%的碳源来自木质纤维素,55.28%的碳源来自非木质纤维素类物质。【结论】金耳生长发育周期内对非木质纤维素(淀粉、蛋白质等)类物质的利用率高于木质纤维素类物质,可从此方面考虑进行培养基配方优化,以提高生物学效率。
关键词: 金耳;碳源;木质纤维素;绝对生物学效率
中图分类号: S567.340.4 文献标志码: A 文章编号:2095-1191(2021)09-2516-08
Regulation on the degradation and transformation of Naematelia aurantialba(Bandoni & M. Zang) Millanes & Wedinmedium
CAO Yao1, LI Rong-chun1*, YANG Lin-lei2, LI Meng-jie2, LUO Xiang-ying2,
SHEN Zhen-hui2, LU Qing-qing2
(1Institute of Edible Fungi, Yunnan Agricultural University, Kunming 650201, China;
2Yunnan Junshijie Biotechnology Co., Ltd., Kunming 650200, China)
Abstract:【Objective】To explore the changes in the degradation and transformation of the medium in different times of growth and development of Naematelia aurantialba(Bandoni & M. Zang) Millanes & Wedin, and to understand the degradation and utilization characteristics of the N. aurantialba for carbon sources. Provided a theoretical basis for the formulation of medium.【Method】The changes in water content, pH, the wet weight and dry weight of the medium, fresh weight of fruitingbody and the dry weight, degradation of lignocellulose were measured in 8 times of growth[0 d(not inoculated, 20, 25, 30, 35, 40, 45, 50 d after vaccination] of the medium, and the changes of various indicators in different growth stages were analyzed. 【Result】In the cycle of N. aurantialba growth and development, water content decreased from 60.27% to 39.00%. At 25 d pH was reduced from 5.50 to 4.55, after which pH was basically stable at about 4.20, with overall acidic conditions. The dry weight of medium from the initial 735.00 g/bag to 530.00 g/bag, the fresh weight of fruitingbody production was 644.71 g/bag. The total medium reduction was 27.89%, of which the biological efficiency was 12.33%, and the absolute respiratory consumption was 15.56%. 30 to 35 d was the first peak of fruitingbody growth, and 40 to 45 d was the second peak of fruiting body growth. In the 0-40 d of N. aurantialba growth and development,respiratory effect with the growth of fruiting body continued to increase. The use of the lignocellulose of the medium parts was in order of cellulose, lignin, hemicellulose andthe utilization rates were 33.90%, 32.79% and 30.76%. The early stage(0 to 30 d) was mainly carbon source of non-lignocellulose substances, and the later stage(30 to 50 d) of growth was mainly carbon source of lignocellulose substances, of which 44.72% of the carbon sources required came from lignocellulose and 55.28% of carbon sources came from non-lignocellulose. 【Conclusion】During the growth and development cycle of N. aurantialba, the utilization rate of non-lignocellulose(starch, protein) substances is proximity to that of lignocellulose. The optimization of medium formulation can be considered from this aspect, in the hope of impro-ving biological efficiency.
Key words: Naematelia aurantialba(Bandoni & M. Zang) Millanes & Wedin; carbon sources; lignocellulose; absolute biological efficiency
Foundation item: National Natural Science Foundation of China-Yunnan Joint Project(U1802231)
0 引言
【研究意义】金耳[Naematelia aurantialba(Bandoni & M. Zang) Millanes & Wedin]在分类学上隶属于真菌界、担子菌门、银耳纲、银耳目、耳包革科、耳包革属,又名脑耳、黄金银耳、金木耳等(Bandoni and Zang,1990;杨林雷等,2020)。金耳鲜嫩子实体口感滑嫩爽口,营养价值相当丰富,富含人体所必需的8种氨基酸及胶质物、碳水化合物、多种维生素和矿物质(汪虹,2005;李元伟,2016)。金耳子实体是异质型子实体,是由金耳菌菌丝和毛韧革菌[Ste-reum hirsutum(Willd) Pers]菌丝组成的复合体,金耳菌丝的生长发育主要依靠毛韧革菌为其提供营养;毛韧革菌具有较强的非纤维素和纤维素物质的分解能力,可提供金耳的生长发育、形成子实体、完成有性生殖过程所需的营养物质,因其生物学特性较为特殊,导致其栽培难度大、菌种制作困难、转化率低、成品率低,因此在栽培中迫切需要了解金耳对栽培基质中不同成分的降解和利用水平,以便筛选最佳培养基配方,实现高产稳产。【前人研究进展】金耳是20世纪我国科技工作者新驯化栽培的珍稀食用菌,金耳的研究主要集中在栽培技术、营养价值及食品开发方面,而营养生理方面研究甚少。金耳的栽培研究为郑淑芳和刘平(1983)首次利用木屑培养基对金耳进行人工栽培,虽获成功,但其出耳率很低;刘平(1985)首次对金耳进行段木栽培,其出耳率为58%;刘正南和郑淑芳(1995)关于有效菌种的提出后,是人工代料栽培金耳高产技术的关键;钟冬季和钟秀媚(2008)对金耳栽培技术研究表明金耳生长发育要经过2个时间段,经过发菌房培养之后到出耳房;田果廷等(2012)对金耳代料栽培技术研究表明,89.8%中药渣、8%玉米粉、1%蔗糖、1%石膏粉、0.2%磷酸二氢钾配方的基质栽培生物学效率达55%;刘书畅等(2019)对金耳菌种生产技术研究表明,原种适宜的恒温培养温度范围为18~22 ℃,变温培养时间为6~9 d,最优原种配方碳氮比为390∶1,其对应配方为木屑91.4%、玉米粉6.6%、石膏1%、糖1%;云南菌视界生物科技有限公司已实现工厂化生产栽培,其现代化栽培菇房于《食药用菌》杂志2020年第4期封面刊登。在金耳的营养价值及食品开发方面,邓超等(2017)研究表明,金耳多糖具有一定的还原能力及较强的清除DPPH和羟基自由基活性,可显著降低四氧嘧啶诱导的糖尿病小鼠血糖含量;康林芝等(2020)利用金耳发酵液作为原料研制出一种新型的食用菌酸奶制品。在金耳营养生理研究方面,回晶等(2007)對金耳液体培养过程中几种胞外酶活性的变化规律研究表明,邻苯二酚氧化酶、愈创木酚氧化酶和漆酶活性高峰均在淀粉酶、羧甲基纤维素酶、半纤维素酶和果胶酶活性降到很低时才出现,说明金耳先利用非木质纤维素类物质后再利用木质纤维素类物质;刘胜贵等(2011)研究纤维素降解真菌的筛选及产酶条件得出金耳能够产生大量的纤维素酶。【本研究切入点】近年来,随着金耳市场需求量不断增加,食用菌大规模人工种植,食用菌生理方面已得到深入研究(高锋等,2017;蔡盼盼等,2019)。研究食用菌基质内营养物质利用特点可对食用菌生长发育进行相应调控,从而提高食用菌产量(袁胜东,2012);而有关金耳基础生理生物学的研究仍较缺乏。【拟解决的关键问题】以金耳工厂化栽培配方为研究对象,测定金耳8个生长阶段栽培基质中含水量、pH、培养料重量、子实体重量和木质纤维素的变化,探索金耳不同生长发育过程中物质的降解与转化规律,初步了解金耳对栽培基质中物质利用顺序、利用率和碳源利用特点等问题,为栽培培养基配方设计及金耳基础生物学研究提供理论依据。
1 材料与方法
1. 1 试验材料
试验所用金耳菌株JSJ-X9为云南菌视界生物科技有限公司保存菌种。培养基配方:棉籽壳30%,木屑56%,麦麸12%,磷酸二氢钾1%,硫酸钙1%,含水量60%。主要仪器和设备:雷磁pH计(PHS-3C,上海仪电科学仪器股份有限公司),恒温恒湿培养箱(HWS-280,宁波江南仪器厂),电热鼓风干燥箱(DHG-9070,上海一恒科学仪器有限公司),电子天平(JA1002,上海浦春计量仪器有限公司),恒温水浴锅(HH-8,金坛市城西春兰实验仪器厂)。
1. 2 试验方法
1. 2. 1 菌包制作及培养 于2020年7月25日—11月初进行试验。按照1.1中培养基配方制作菌包,于聚丙烯菌袋(15 cm×55 cm)中装入均匀搅拌后的培养料重2.00 kg(湿重)。在灭菌条件为121 ℃、0.103 MPa下高温高压蒸汽灭菌90 min。灭菌后在无菌条件下再次称重,保持菌包重量一致,将准备接种于菌包的菌种进行称重。为避免试验中不必要的误差,需要保持菌种重量一致、菌包重量一致,使所测指标数据更可靠。接种后使用美纹纸进行封口,置于18 ℃下黑暗培养发菌,湿度50%,待菌落直径达7 cm时揭去美纹纸,调整菇房管理参数为湿度90%、光照800 Lux(8 h/d)、CO2浓度≤800 ppm,培养至不同时间段后进行取样。
1. 2. 2 取样 分为8个时间进行菌包取样,时间分别为未接种(0 d),以及接种后20、25、30、35、40、45和50 d。从工厂化栽培的情况来看,在培养至20 d时,菌丝刚好长满菌包,设定从此时开始取样,此后每隔5 d取1次样,直至成熟采收。每个指标3个重复,一袋菌包则视为1个重复。
1. 2. 3 测定指标及方法 通过雷磁pH计测定菌包pH;称重菌包培养料湿重、每袋菌包子实体总鲜重及总干重。纤维素、半纤维素和木质素的检测方法参照王玉万和徐文玉(1987)的方法,略加改进。根据上述数据计算:含水量(%)=(培养料烘干前重量-培养料烘干后重量)/培养料烘干前重量×100;培养料干重(g)=培养料湿重×(1-含水量);总培养基失重(g)=培养料初始干重-培养料最终干重;总培养基失重率(%)=(培养料初始干重-培养料最终干重)/培养料初始干重×100;子实体百分比(%)=子实体各阶段重量/子实体最终重量×100;培养料阶段干重百分比(%)=培养料各阶段干重/培养料最初干重×100;各物质减少量(g)=初始量-各阶段量;
参照倪新江等(2001)的方法计算:绝对生物学效率(%)=子实体干重/总培养料干重×100。
参照初洋等(2014a)的方法计算:木质纤维素组分减少量(g)=纤维素减少量+半纤维素减少量+木质素减少量;木质纤维素各组分利用率(%)=各物質减少量/各物质初始量×100;呼吸消耗率(%)=总培养基失重率-绝对生物学效率;木质纤维素各组分与培养基失重比值(%)=各物质所测利用量/总培养基失重×100。
1. 3 统计分析
试验数据使用Excel 2019记录及数据分析,运用SPSS 25.0进行均值分析、方差分析及单因素ANOVA检验。
2 结果与分析
2. 1 金耳不同生长时间含水量及pH变化
在金耳生长发育过程中,水分是反应活动的底物,调控各生命活动。由表1可知,随着金耳子实体的生长发育,菌包含水量不断下降,由最初的60.27%降至最终的39.00%。30~35 d和40~45 d含水量降速快,表明这2个时间段内生理生化活动加强,金耳生长较为旺盛,导致含水量减少,35、40和45 d的含水量与其他生长时间差异显著(P<0.05,下同)。金耳子实体生长发育周期内pH总体较稳定,0~20 d主要为毛韧革菌菌丝生长,金耳子实体生长不明显;当毛韧革菌菌丝长满培养料,pH有所下降,说明毛韧革菌菌丝长满培养料的过程中分泌了大量的胞外酶来降解基质内大分子营养物质,基质内代谢旺盛,引起基质中离子浓度变化;25 d时pH由最初的5.50下降至4.55,30~50 d的pH无显著差异(P>0.05,下同),pH维持在4.20左右。因此,金耳的生长发育是在酸性条件下进行的。
2. 2 金耳不同生长时间培养料重量变化
由表1可知,在金耳生长发育周期内,培养料干重由735.00 g/袋降至530.00 g/袋,平均每袋降解205.00 g干物质,培养基失重率27.89%(干重)。生长20 d后培养料干重减少6 g/袋,20~25 d减少10.00 g/袋,25~30 d减少22.00 g/袋,30~35 d减少69.00 g/袋,35~40 d时减少46.00 g/袋,40~45 d减少52.00 g/袋,45~50 d未减少,表明金耳的生长已趋于成熟。30~35 d培养料干重降解速度最快,表明此阶段为金耳生长发育的第1次高峰期;35~40 d干重减少量相对于30~35 d有所降低,可能与环境条件管理有关,环境中的温度、湿度或CO2浓度不利于金耳的生长发育;40~45 d为金耳生长发育的第2次高峰期;45~50 d物质降解量达最低。从降解量来看,培养基内剩余近72.00%干物质未利用,可考虑进一步优化培养基的重量来达到物质的最佳利用率。
2. 3 金耳不同生长时间子实体重量变化
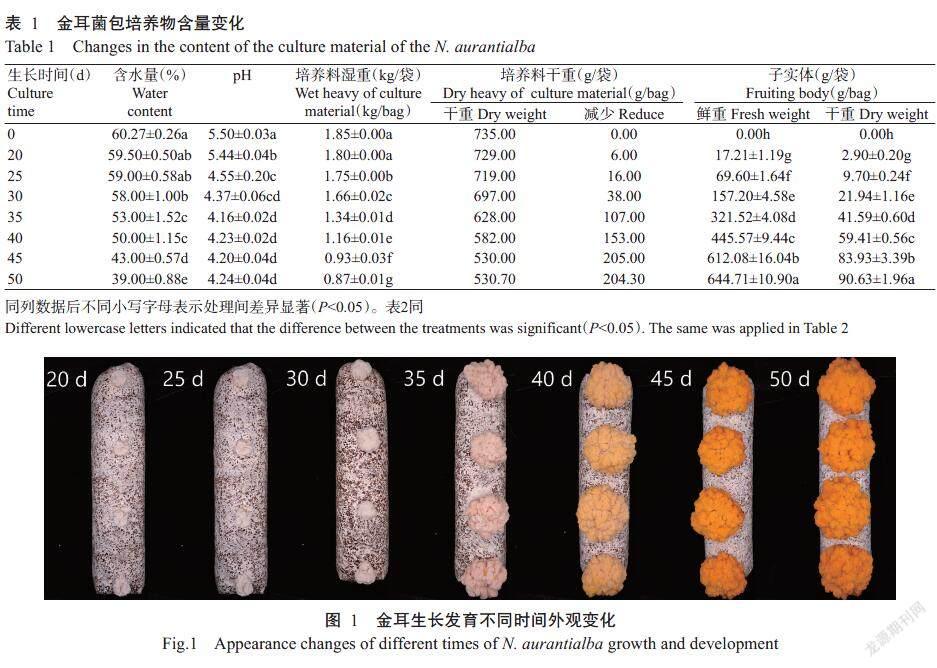
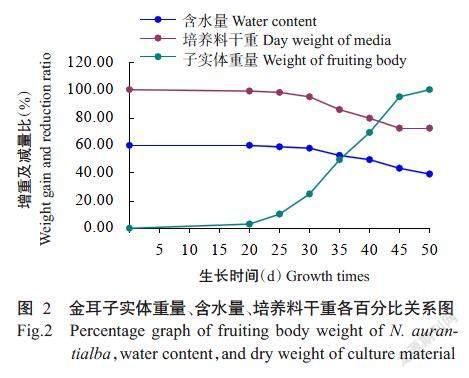
由表1可知,在不同培养时间,金耳子实体生长发育均达显著差异。子实体鲜重产量为644.71 g/袋,干重为90.63 g/袋,绝对生物学效率12.33%。培养时间30~35 d子实体生长最快,重量以倍数增加;35~40 d子实体的重量增长值小于30~35 d;40~45 d子实体增长量又以倍数增加,子实体重量出现2个增重高峰期(30~35和40~45 d);50 d时子实体增重量相对20~45 d各阶段来说达最低值,培养料干重于此时间段未发生减重。从外观看子实体已成熟,开始释放孢子,表明此时子实体吸收、积累营养物质的能力已达到饱和状态,生长发育已停止,可进行采收(图1)。子实体增重量与培养料干重、含水量变化呈负相关,子实体不断的增长,含水量及培养料干重不断下降(图2)。
2. 4 金耳不同生长阶段呼吸作用消耗分析
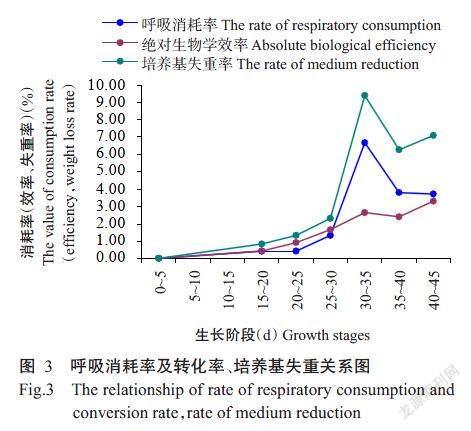
由表1中培养料、子实体干重数据可计算出0~20、20~25、25~30、30~35、35~40和40~45 d各阶段的呼吸消耗率,依次为0.42%、0.44%、1.33%、6.71%、3.83%和3.74%。在40 d前呼吸作用消耗率与绝对生物学效率、培养基失重率呈现出了相应的变化趋势,随着绝对生物学效率、培养基失重率的增加而增加,于30~35 d出现了1次高峰,35~40 d下降,在45 d时呼吸消耗率发生降低,未伴随上升(图3)。呼吸消耗率与子实体绝对生物学效率、培养基失重率在0~40 d呈正相关,呼吸作用越强,金耳子实体生长越快,此时培养基失重率也相应上升。由2.2及2.3可知0~50 d总培养基失重率27.89%,绝对生物学效率12.33%,所以子实体总呼吸作用消耗率为15.56%,总培养料干重为735.00 g/袋,表明在整个生长发育周期内子实体呼吸作用消耗了114.37 g/袋基质,作为呼吸作用消耗损失,以CO2、能量的形式散发到空气中。
2. 5 金耳不同生长时间木质纤维素的降解利用
由表2可知,在金耳子实体生长发育至25 d时,木质纤维素组分中只有纤维素物质发生了减少,纤维素含量在0与25 d时具有显著性差异,而半纤维素与木质素的含量无显著性差异。各物质减少量在45 d时达最大值,为吸收营养的顶峰。生长发育至50 d时,物质减少量未发生变化,纤维素、半纤维素和木质素含量在45与50 d均无显著性差异。0~50 d木质纤维素组分利用顺序及利用率依次为纤维素(33.90%)、木质素(32.79%)、半纤维素(30.76%)。如图4所示,木质纤维素组分的降解利用在随着培养时间增加而不断上升,30~35 d各物质利用率相比0~30 d大幅度上升,物质降解增多,且结合子实体重量变化数据,可知该阶段生长发育急剧加快,表明需要的营养则较多,此生长时期的环境条件应谨慎管理。
2. 6 金耳不同生长阶段木质纤维素降解量与培养基失重的比值
菌包在培养0~20 d内主要为毛韧革菌菌丝生长期,金耳子实体膨大较为缓慢。由表3可知,木质纤维素组分中,纤维素降解2.17 g,培养基失重6.00 g,木质纤维素降解量与培养基失重的比值为36.17%,即36.18%的营养由木质纤维素提供,另外63.83%的营养物质由非木质纤维素提供,表明主要利用的是非木质纤维素组分物质(淀粉类)为碳源。30~35、35~40和40~45 d为金耳子实体吸收营养的高峰期,木质纤维素与培养基失重比值为50.29%、50.10%和49.02%,木质纤维素与培养基的比值均基本达到或超过50.00%,主要利用菌包培养物中的木质纤维素组分作为碳源。因此,生长前期(0~30 d)主要是以非木质纤维素类物质作为碳源,生长后期(30~50 d)主要以非物质纤维素类物质作为碳源。整个生长发育周期内,木质纤维素降解量为91.68 g,培养基失重205.00 g,二者比值为44.72%。说明,金耳子实体生长发育所需要的44.72%的碳源来自木质纤维素,另外55.28%的碳源则来自非木质纤维素类物质。
3 讨论
相关研究表明菌体的呼吸强度与绝对生物学效率有一定的正相关性(彭强,2007);本研究中,金耳生长发育过程中,呼吸消耗与绝对生物学效率在0~40 d生长时间内呈正相关。生长发育过程中30~35与40~45 d金耳子实体重量以倍数明显增加,这2个阶段分别为金耳生长的第1和第2个高峰期,应当增加菇房的通风换气,谨慎管理菇房环境条件,以利于菌丝生长情况与子实体生长发育,从而可进一步提高生物学效率。
大多数食用菌为腐生性真菌,自身不能合成营养物质,需从栽培基质中分解各种碳、氮源等大分子物质。菌包中菌丝不能直接对栽培基质中的营养进行吸收,而是通过自身分泌的胞外酶对大分子物质进行降解成为可吸收的小分子物质。随着食用菌大规模人工种植,食用菌的营养生理也得到了多方面的研究,其中以香菇的研究(冯志勇等,2000;張权,2016;陈小敏等,2019)较为广泛。通过对食用菌营养生理进行研究可开发新的栽培配方以及技术。金耳为金耳菌与毛韧革菌2个物种混合生长,其生物学特性较为复杂。单独金耳菌几乎没有独立生长的能力,不能提供自身营养,而是靠毛韧革菌分解栽培基质来提供营养。本研究中,木质纤维素组分利用顺序及最大利用率依次为纤维素>木质素>半纤维素;从最大利用率数据来看,最大利用率仅38.00%,这一数值较不可观。后续研究应当优化培养基配方、培养料重量来增加各物质的利用率,从而进一步达到高产、稳产。金耳整个生长发育阶段55.28%的碳源来自非木质纤维素,44.72%的碳源来自木质纤维素,说明金耳对木质纤维素物质利用率比非木质纤维素物质(淀粉、蛋白质)低。在对培养基配方进行优化时,可尝试适当提高含淀粉、蛋白质类物质的配比,可能会取得较好的生物学效率。本研究得出菌包中木质纤维素各物质利用率均未达50.00%,菌渣中所剩各物质含量还可达50.00%,且从培养基干重数据得到总培养基失重率仅27.89%。可见,金耳所剩菌渣具有再利用价值,可考虑如何有效地进行二次出菇或利用其栽培一些降解木质纤维素能力强的菇,可能会取得好的生物学效率,也可作为其他作物种植的肥料。如利用杏鲍菇菌渣栽培玉木耳和鲍鱼菇,杏鲍菇菌渣添加量为20%可提高玉木耳的产量(钟方翼和曾凡清,2020);在菌渣添加比例为30%时鲍鱼菇产量最高,生物学效率达77.48%(杭中桥等,2017)。
在回晶等(2007)对金耳液体培养的研究中,通过酶活角度得到了金耳生长前期主要是利用淀粉类物质作为碳源,本研究结果与其相同,金耳在生长前期(0~30 d)主要是以非木质纤维素类物质作为碳源,生长后期(30~50 d)主要是以木质纤维素类物质作为碳源;金耳利用碳源的这一规律与鸡腿菇(倪新江等,2002)、杏鲍菇(初洋等,2014b)相似。下一步的研究工作也将会从酶活角度来对金耳的生长变化过程进行更深入的探索。另外,本研究结果与培养基配方及菇房环境管理等有着重大关系,若培养基配方不同,则会导致结果有较大差异。
4 结论
金耳生长发育周期内对非木质纤维素(淀粉、蛋白质等)类物质利用率高于木质纤维素类物质,可从此方面考虑进行培养基配方优化,以提高生物学效率。
参考文献:
蔡盼盼,张文强,张昊琳,王清辉,张国庆,陈青君. 2019. 双孢蘑菇培养料理化指标及酶活与其产量相关的多重分析[J]. 农业工程学报,35(7):231-237. [Cai P P,Zhang W Q,Zhang H L,Wang Q H,Zhang G Q,Chen Q J. 2019. Multiple analysis of relationship between yield of Agaricus bisporus with physicochemical properties and enzyme activity of substrates[J]. Transactions of the Chinese Society of Agricultural Engineering,35(7):231-237.] doi:10.11975/j.issn.1002-6819.2019.07.029.
陈小敏,吴海冰,向泉桔,曾先富,张小平,辜运富. 2019. 不同温度培养对香菇漆酶活性及转录表达的影响[J]. 四川大学学报(自然科学版),56(1):155-160. [Chen X M,Wu H B,Xiang Q J,Zeng X F,Zhang X P,Gu Y F. 2019. Effects of different temperature cultures on laccase activity and transcriptional expression of Letinous edodes[J]. Journal of Sichuan University(Natural Science Edition),56(1):155-160.] doi:10.3969/j.issn.0490-6756. 2019.01.027.
初洋,董新伟,倪新江,赵金良. 2014a. 洛巴伊大口蘑对栽培基质的降解及其相关胞外酶活性变化[J]. 北方园艺,(6):147-149. [Chu Y,Dong X W,Ni X J,Zhao J L. 2014a. Degradation of the cottonseed shell medium and activity changes of related extracellular enzymes from Tricholoma lobayense at different stages of development[J]. Northern Horticulture,(6):147-149.]
初洋,董新伟,倪新江,赵金良. 2014b. 杏鲍菇对栽培基质的降解与转化[J]. 北方园艺,(15):158-160. [Chu Y,Dong X W,Ni X J,Zhao J L. 2014b. Degradation and transformation of the cottonseed shell medium by Pleurotus eryngii[J]. Northern Horticulture,(15):158-160.]
邓超,付海田,尚京迎,乔祉琦,赵楚,陈敬华. 2017. 金耳发酵液多糖的制备、分析及生物活性研究[J]. 食品与生物技术学报,36(1):67-73. [Deng C,Fu H T,Shang J Y,Qiao Z Q,Zhao C,Chen J H. 2017. Preparation,chemical analysis and biological activity of polysaccharides from fermented Tremella aurantialba[J]. Journal of Food Science and Biotechnology,36(1):67-73.] doi:10.3969/j.issn.1673-1689.2017.01.012.
馮志勇,潘迎捷,陈明杰,谭琦,贺冬梅. 2000. 香菇发育生理研究进展[J]. 食用菌学报,7(4):53-60. [Feng Z Y,Pan Y J,Chen M J,Tan Q,He D M. 2000. Recent studies on the developmental physiology of Lentinula edodes[J]. Acta Edulis Fungi,7(4):53-60.] doi:10.16488/j.cnki. 1005-9873.2000.04.012.
高锋,曹旸,何明霞,刘静,方艺伟,张春霞. 2017. 香菇、杏鲍菇与平菇部分胞外酶种类及活性变化规律的对比分析[J]. 北方园艺,(23):199-206. [Gao F,Cao Y,He M X,Liu J,Fang Y W,Zhang C X. 2017. Type and activity variation of extracellular enzymes of Lentinus edodes,Pleurotus eryngii and Pleurotus ostreatus[J]. Northern Horticulture,(23):199-206.] doi:10.11937/bfyy.20171421.
杭中桥,杨鹏,王建设,宫志远,万鲁长,韩建东. 2017. 利用杏鲍菇菌渣栽培鲍鱼菇试验[J]. 中国食用菌,36(3):33-36. [Hang Z Q,Yang P,Wang J S,Gong Z Y,Wan L C,Han J D. 2017. Cultivation of Pleurotus abalonus using Pleurotus eryngii spent substrate[J]. Edible Fungi of China,36(3):33-36.] doi:10.13629/j.cnki.53-1054.2017.03. 008.
回晶,赵文静,邹志远,张月. 2007. 金耳液体培养过程中几种胞外酶活性的变化规律[J]. 食用菌学报,14(3):29-32. [Hui J,Zhao W J,Zou Z Y,Zhang Y. 2007. Changes in extracellular enzyme activities during submerged culture of Tremella aurantialba[J]. Acta Edulis Fungi,14(3):29-32.] doi:10.3969/j.issn.1005-9873.2007.03.005.
康林芝,吴居雄,唐惠妍,门戈阳,刘主. 2020. 金耳发酵液酸奶制作及品质分析研究[J]. 食品研究与开发,41(11):141-146. [Kang L Z,Wu J X,Tang H Y,Men G Y,Liu Z. 2020. Study on liquid fermentation of Tremella aurantialba yoghurt and quality analysis[J]. Food Research and Development,41(11):141-146.] doi:10.12161/j.issn. 1005-6521.2020.11.024.
李元伟. 2016. 金耳子实体化学成分及其体外抗氧化活性研究[D]. 长春:吉林农业大学. [Li Y W. 2016. Studies on chemical composition and antioxidant activity in vitro from Tremella aurantialba fruiting bodies[D]. Changchun:Jilin Agricultural University.]
劉平. 1985. 金耳人工段木栽培研究初报[J]. 中国食用菌,(2):6-8. [Liu P. 1985. The study of wood cultivation in the artificial section of Tremella aurantialba was first reported[J]. Edible Fungi of China,(2):6-8.]
刘书畅,李荣春,马布平,罗祥英,李梦杰,曹瑶,闻邵峰,周忠发,杨林雷. 2019. 金耳菌种生产技术研究[J]. 中国食用菌,38(9):94-99. [Liu S C,Li R C,Ma B P,Luo X Y,Li M J,Cao Y,Wen S F,Zhou Z F,Yang L L. 2019. Research on the production technology of Tremella aurantialba bacteria[J]. Edible Fungi of China,38(9):94-99.] doi:10.13629/j.cnki.53-1054.2019.09.021.
刘胜贵,严明,邹娟,吴镝,胡兴. 2011. 纤维素降解真菌的筛选及其产酶条件[J]. 中国农学通报,27(7):102-106. [Liu S G,Yan M,Zou J,Wu D,Hu X. 2011. The scree-ning of biodegrading cellulose fungi and its cellulase-producing conditions[J]. Chinese Agricultural Science Bulletin,27(7):102-106.]
刘正南,郑淑芳. 1995. 金耳的生理特性及有效优良菌种的制备原理[J]. 中国食用菌,14(5):10-11. [Liu Z N,Zheng S F. 1995. The manufacturing principles of the effective fine species and its physiological nature of Tremella aurantialba[J]. Edible Fungi of China,14(5):10-11.]
倪新江,梁丽琨,丁立孝,初洋,鞠宝,潘迎捷. 2001. 巴西蘑菇对木质纤维素的降解与转化[J]. 菌物系统,(4):526-530. [Ni X J,Liang L K,Ding L X,Chu Y,Ju B,Pan Y J. 2001. Degradation and transformation of lignocellulose by Agaricus blazei[J]. Mycosystema,(4):526-530.] doi:10.3969/j.issn.1672-6472.2001.04.016.
倪新江,梁丽琨,由翠荣,冯志勇,潘迎捷. 2002. 鸡腿菇对棉籽壳的降解与转化[J]. 微生物学通报,29(2):1-4. [Ni X J,Liang L K,You C R,Feng Z Y,Pan Y J. 2002. Degradation and transformation of cotton seed hulls by Coprinus Comatus[J]. Microbiology China,29(2):1-4.] doi:10.3969/j.issn.0253-2654.2002.02.001.
彭强. 2007. 黑木耳培养料及营养生理特性研究[D]. 哈尔滨:东北农业大学. [Peng Q. 2007. Studies on substrates and nutrient physiological characteristics of Auricularia auricula[D]. Harbin:Northeast Agricultural University.] doi:10.7666/d.y1165146.
田果廷,陈卫民,苏开美,柴红梅,李树红,赵永昌. 2012. 金耳代料栽培技术研究[J]. 食用菌学报,19(1):43-46. [Tian G T,Chen W M,Su K M,Chai H M,Li S H,Zhao Y C. 2012. Cultivation of Tremella aurantialba using different compost formulae[J]. Acta Edulis Fungi,19(1):43-46.] doi:10.16488/j.cnki.1005-9873.2012.01.011.
汪虹. 2005. 金耳药理活性及其多糖结构研究进展[J]. 食用菌学报,12(4):53-56. [Wang H. 2005. Advances in the study on the pharmacology and structure analysis of polysaccharide of Tremella aurantialba[J]. Acta Edulis Fungi,12(4):53-56.] doi:10.3969/j.issn.1005-9873.2005.04. 012.
王玉万,徐文玉. 1987. 木质纤维素固体基质发酵物中半纤维素、纤维素和木质素的定量分析程序[J]. 微生物学通报,14(2):81-84. [Wang Y W,Xu W Y. 1987. Quantitative analysis procedures for hemicellulose,cellulose and lignin in lignocellulose solid substation fermentations[J]. Microbiology China,14(2):81-84.] doi:10.13344/j.microbiol.china.1987.02.012.
杨林雷,李荣春,曹瑶,李梦杰,罗祥英,杨晓君,闻绍峰,沈真辉,陆青青,子灵山. 2020. 金耳的学名及分类地位考证[J]. 食药用菌,28(4):252-255. [Yang L L,Li R C,Cao Y,Li M J,Luo X Y,Yang X J,Wen S F,Shen Z H,Lu Q Q,Zi L S. 2020. Research on the scientific name and taxonomic status of “Jin Er”[J]. Edible and Medicinal Mushrooms,28(4):252-255.]
袁胜东. 2012. 猴头菌生长过程中基质内营养物质含量和胞外酶活研究[D]. 雅安:四川农业大学. [Yuan S D. 2012. Study of nutrient content and extracellular enzyme activity in substrate during Hericium erinaceus growth[D]. Yaan:Sichuan Agricultural University.]
张权. 2016. 香菇胞外酶活性变化规律和农艺性状研究[D]. 新乡:河南科技学院. [Zhang Q. 2016. Study on the change regularity of extracellular enzyme activity and agronomy characters of Lentinula edodes[D]. Xinxiang:Henan Institute of Science and Technology.]
郑淑芳,刘平. 1983. 金耳人工栽培研究初报[J]. 食用菌科技,(1):15-16. [Zheng S F,Liu P. 1983. The first report of the study of artificial cultivation of Tremella aurantialba[J]. Edible Fungi of China,(1):15-16.]
鐘冬季,钟秀媚. 2008. 金耳栽培技术[J]. 食用菌,(4):50-51. [Zhong D J,Zhong X M. 2008. Tremella aurantialba cultivation technology[J]. Edible Fungi,(4):50-51.] doi:10.3969/j.issn.1000-8357.2008.04.036.
钟方翼,曾凡清. 2020. 杏鲍菇菌渣对玉木耳栽培的影响[J]. 中国食用菌,39(6):6-7. [Zhong F Y,Zeng F Q. 2020. Effects of Pleurotus eryngii chaff on Auricularia cornea cultivation[J]. Edible Fungi of China,39(6):6-7.] doi:10. 13629/j.cnki.53-1054.2020.06.002.
Bandoni R J,Zang M. 1990. On an undescribed Tremella from China[J]. Mycologia,82(2):270-273. doi:10.1080/0027 5514.1990.12025876.
(责任编辑 邓慧灵)
