ACE抑制肽生物信息及核桃蛋白虚拟酶切研究
2021-09-12史珂雨赵慧琳李明鑫秦兆盈吴迪杜明王聪
史珂雨,赵慧琳,李明鑫,秦兆盈,吴迪,杜明,王聪
(1.大连工业大学食品学院,辽宁 大连 116034;2.大连工业大学实验仪器中心,辽宁 大连 116034)
目前,人工合成的降血压药物有200余种,但需要终身口服,而且大多存在引起低血压、咳嗽、血钾升高、胎儿畸形等副作用。因此,急需开发安全性更高的食源性血管紧张素转换酶(angiotensin I-converting enzyme,ACE)抑制肽。
核桃具有很高的食用价值和药用价值,在我国中医药学中有着广泛的应用。核桃仁营养丰富,含有16%~19%的优质蛋白质[1]。以核桃为基料的ACE抑制肽研究,在酶法水解工艺筛选、分离纯化条件优化等方面已经积累了比较丰富的经验,在结构鉴定和构效关系方面也开始进行逐步深入的探讨。随着科技飞速进步,将新兴的计算机生物信息技术与传统多肽研究方法相结合的探索,即将展现出特有的优势。
生物信息学方法主要依赖于大量试验数据建立的数据库,以此为基础对目标蛋白质的各种生物信息、活性功能、分子间作用机理进行分析判断,可以减少生物制备纯化鉴定的繁琐工序和制备成本,加快试验进程[2-5]。随着ACE抑制肽的广泛深入研究,蛋白一级序列数据库的逐步建立,利用计算机模拟复合酶水解蛋白产生ACE抑制肽的这种思路,对于实践操作具有较好的指导作用。利用生物信息技术阐述ACE抑制肽分子特性,尤其是氨基酸在其中的分布规律,分析核桃蛋白酶解产物中降血压肽的分布情况,对于ACE抑制肽的分离纯化及构效关系研究具有重要的指导作用,对于核桃降血压肽的深度开发具有重要的应用价值。
1 材料与方法
1.1 ACE抑制肽分子信息库建立
ACE抑制肽的分子信息收集主要来自于两部分,绝大部分从BIOPEP数据库中获得[6],另外一部分从Web of Science等文献数据库中通过文献检索获得。
1.2 ACE抑制肽的分子特性与结构分析
主要关注ACE抑制肽的几方面信息:分子量、等电点、表面净电荷、疏水性、N-末端氨基酸、C-末端氨基酸等,建立这些分子信息与ACE抑制活性之间的内在规律性联系。为后续ACE抑制肽的分离制备、活性分析及构效关系研究提供一定的理论参考。
1.3 虚拟酶切数据库选择
采用PeptideCutter软件数据库进行虚拟酶切位点分析,该数据库的互联网地址为https://web.expasy.org/peptide_cutter/[7]。
1.4 核桃蛋白虚拟酶切位点分析
核桃种子储藏蛋白(GeneBank:AAW29810.1)分子由507个氨基酸组成,序列从美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)蛋白数据库(http://www.ncbi.nlm.nih.gov/ncbisearch)中获得[8]。采用PeptideCutter进行虚拟酶切位点分析,主要选取胃肠消化道中常见的蛋白酶。通过酶切位点分析推测其酶切产物,进而将这些虚拟酶切产物的氨基酸序列作为产物活性判定的依据之一。
2 结果与分析
2.1 ACE抑制肽分子组成与活性关系分析
本研究经过搜索数据库以及文献总结归纳,选取了目前报道的691条具有ACE抑制活性的肽段,针对这些ACE抑制肽的分子结构特点展开分析,建立了这些分子信息与ACE抑制活性之间的内在规律性联系。
ACE抑制肽的肽链长度和分子量的分布情况见图1。
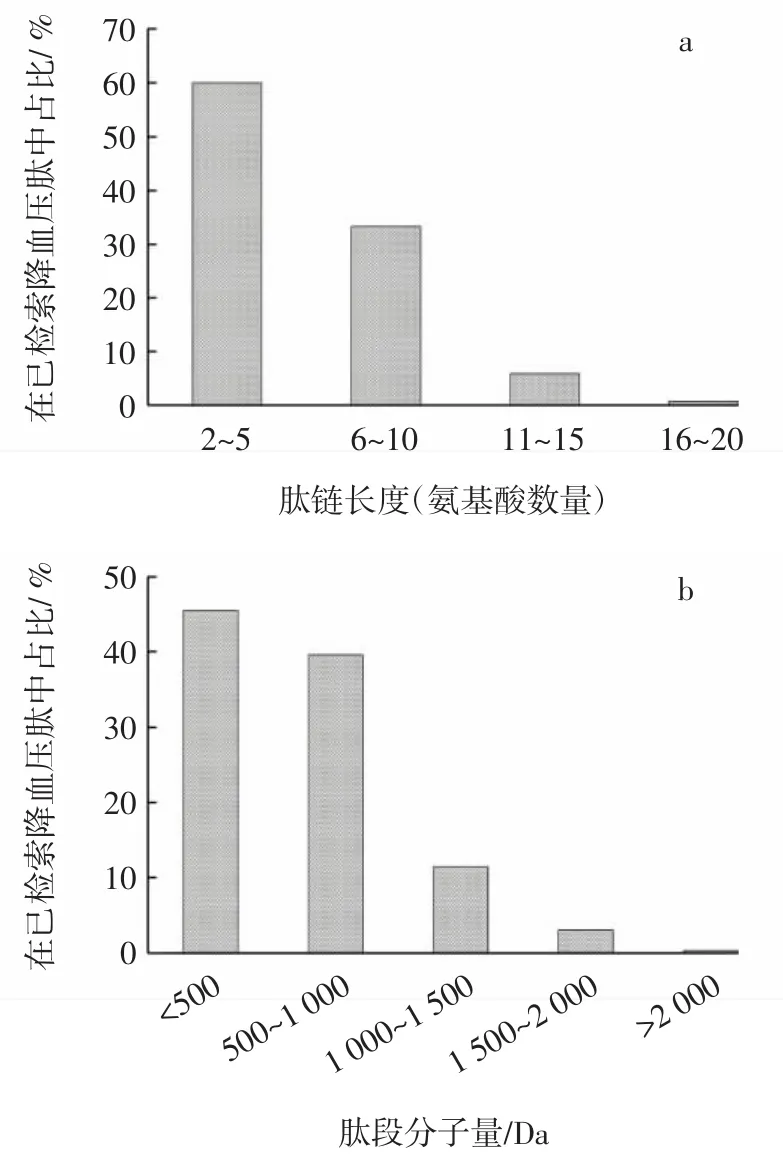
图1 ACE抑制肽的肽链长度和分子量的分布情况Fig.1 Relationship of peptide chain length and ACE activity
研究显示ACE抑制肽的降压活性与其相对分子质量大小、肽链长度有关。本研究发现(如图1),ACE抑制肽通常含有2个~20个氨基酸。由图1a可知,长度在2个~5个氨基酸的肽有415条、6个~10个氨基酸的肽有230条、10个~15个氨基酸的肽有41条、16个~20个氨基酸的肽只有5条,分别占到所汇总ACE抑制肽总数的60.06%、33.28%、5.93%和0.72%。肽链含有2个~5个氨基酸的短肽出现的频率最高。肽的分子量与肽链长度成正比,综合考虑大部分肽的纯化鉴定等研究中,均与分子量直接关联,因此也应该针对分子量进行分析。
由图1b可知,分子量分布在500 Da以下的肽有315条、500 Da~1 000 Da的肽有274条、1 000 Da~1 500 Da的肽有79条、1 500 Da~2 000 Da的肽有21条、而分子量大于2 000 Da的肽仅有2条,分别占到所汇总ACE抑制肽总数的45.58%、39.65%、11.43%、3.04%和0.29%。从统计学的规律来看,分子量小于1 000 Da的ACE抑制肽所占比例较大,其中分子量500 Da以下的ACE抑制肽比例最大。这说明小分子肽更容易进入到ACE活性中心,发挥抑制作用。而肽链长度过长,影响活性肽进入到ACE三级结构的内部,可能由于空间位阻效应,不利于与ACE活性位点结合。
2.2 ACE抑制肽末端氨基酸与活性关系分析
ACE抑制肽的末端氨基酸分布情况见图2。
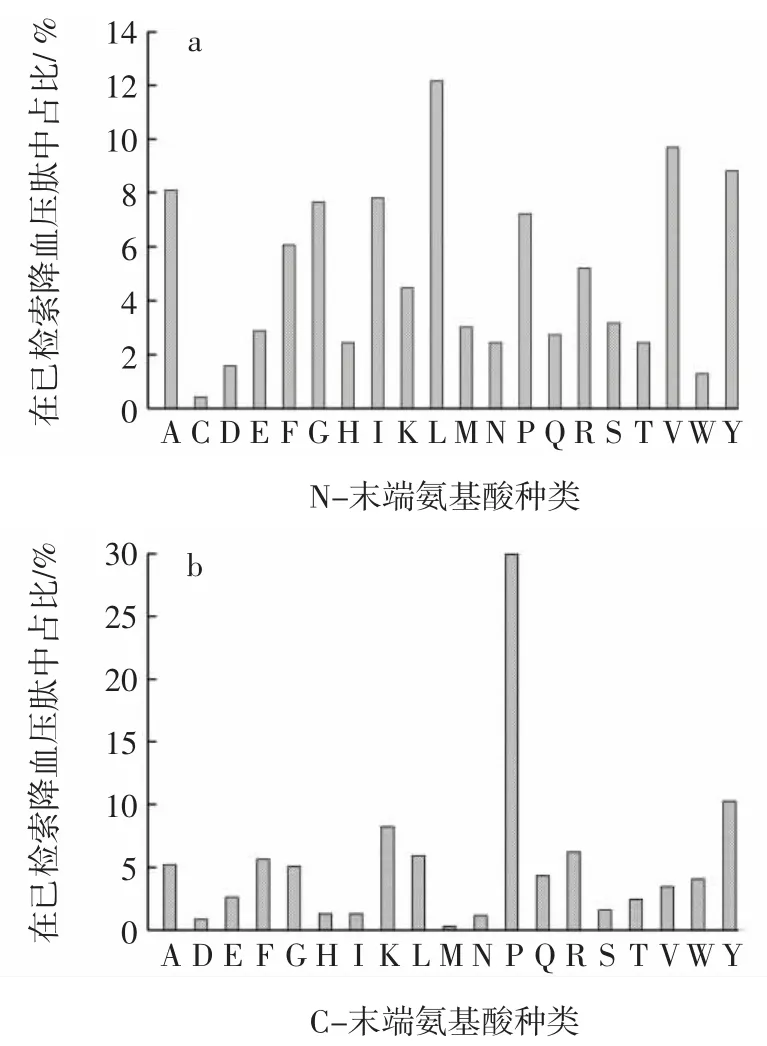
图2 ACE抑制肽的末端氨基酸分布情况Fig.2 Relationship of amino acid properties of peptides and ACE activity
目前,ACE抑制肽的结构和功能的关系还没有彻底阐明,但是研究普遍认为,ACE抑制肽的抑制活性和其自身的结构和氨基酸顺序有关。由图2a可知,对ACE抑制肽的N-末端氨基酸分布情况进行统计分析,发现有8种氨基酸在ACE抑制肽的N-末端出现的频率较高,它们分别为 Leu、Val、Tyr、Ala、Ile、Gly、Pro和Phe。其中Leu出现频率最高为12.15%,其余7种氨基酸的出现频率分别为9.69%、8.83%、8.10%、7.81%、7.67%、7.23%和6.08%。同时,从中也发现Asp、Trp和Cys在ACE抑制肽的N-末端出现频率很低,分别为1.59%、1.30%、0.43%,说明这些氨基酸在肽链的N-末端时,其降血压活性可能处在较低水平。
由图2b可知,对691种ACE抑制肽的C-末端氨基酸进行分析研究,Pro表现出明显的优势,在C-末端出现频率达到29.96%。此外,还有7种主要氨基酸在ACE抑制肽的C-末端出现的频率较高,它们分别为Tyr、Lys、Arg、Leu、Phe、Ala 和 Gly,对应出现频率分别为 10.27% 、8.25% 、6.22% 、5.93% 、5.64% 、5.21% 和5.06%,主要为芳香族氨基酸(Tyr、Phe)、带正电荷的碱性氨基酸(Lys、Arg)或侧链脂肪烃氨基酸(Leu、Ala和Gly),同时,从图中也发现 Met、Asp、Asn、Ile 和 His在ACE抑制肽的C-末端出现频率很低,对应出现频率分别为0.29%、0.87%、1.16%、1.30%和1.30%,而Cys在C-末端甚至没有出现,说明这些氨基酸在肽链的C-末端时,其降血压活性可能不高。
2.3 ACE抑制肽分子表面电荷与活性关系分析
ACE抑制肽的等电点及净电荷分布情况见图3。
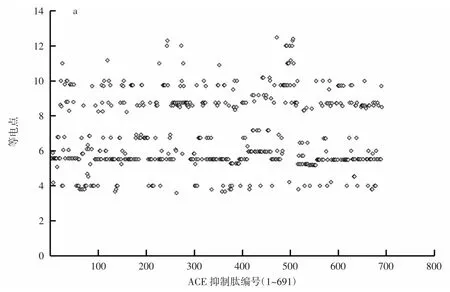

图3 ACE抑制肽的等电点及净电荷分布情况Fig.3 Relationship of pI and charge of peptides and ACE activity
在对ACE抑制肽分子特性与构效关系的研究中,关于肽的等电点和净电荷对ACE抑制活性的作用效果的探讨很少见。由图3a可以看出,等电点在pH 7以下的肽共有460条,占所统计肽总数的66.57%;等电点在pH 8以上的肽共有224条,占所统计肽总数的32.42%;等电点在pH 7~8即生理pH左右的肽仅有7条,仅占所统计肽总数的1.01%。由图3b可以看出,691种ACE抑制肽的净电荷规律以不带电或者带一个正电荷为主,带正电荷的肽有286条,占所统计肽总数的41.39%;不带电荷的肽有325条,占所统计肽总数的47.03%;而带有负电荷的肽仅有80条,占所统计肽总数的11.58%。因此推测负电荷对抑制活性的促进作用不明显。总体来看,不带电或带有少量正电荷更有利于发挥ACE抑制活性。
ACE抑制肽的等电点数值基本集中在pH 5~7和pH 8~10两个范围内,分别占比55.86%和28.94%。在pH 7.3左右的范围内,具有活性的ACE抑制肽非常少。分析这种情况可知,活性肽在等电点附近会聚集变性,不能与ACE分子顺利结合而失去生理活性,而等电点pH5~7或pH8~10的ACE抑制肽则可以在生理pH条件下带正电荷或者负电荷,有带电荷和羧酸基团的肽与ACE更容易形成氢键[9]。这样更容易与ACE分子之间形成氢键或者净电荷相互作用,有利于肽与ACE之间形成稳定的复合结构。
2.4 ACE抑制肽疏水性与活性关系分析
ACE抑制肽的亲水指数和亲水氨基酸分布情况见图4。
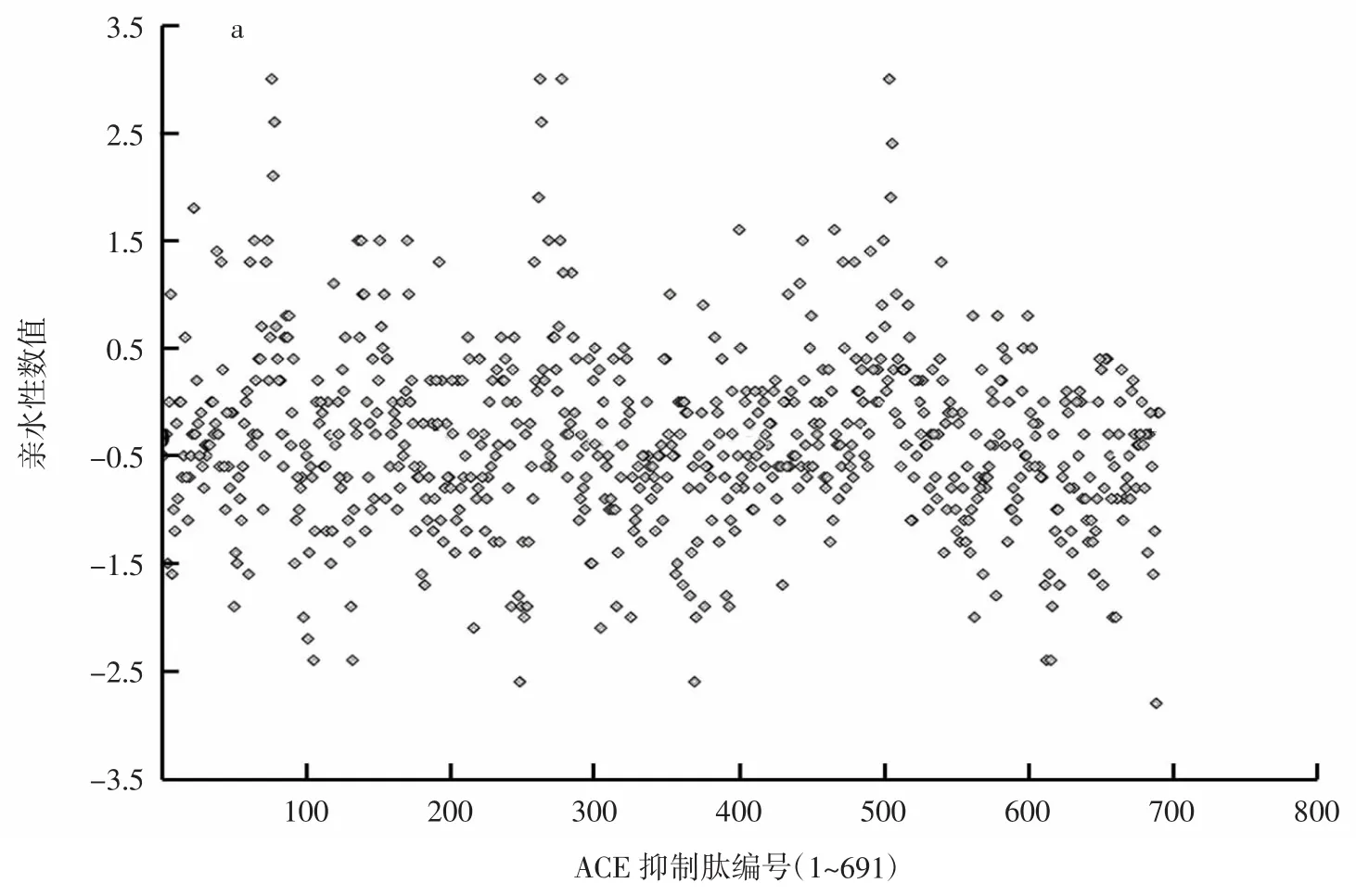
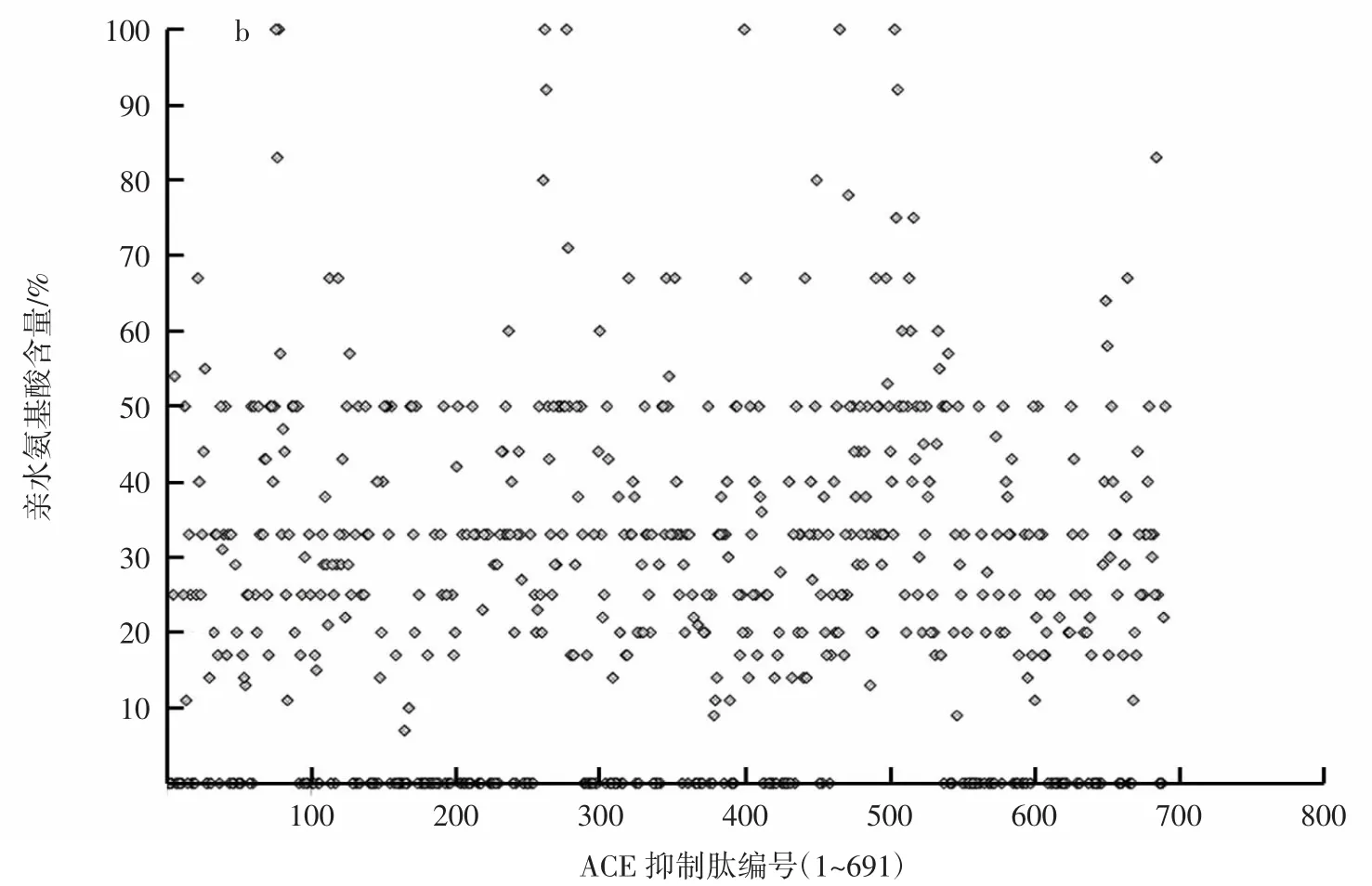
图4 ACE抑制肽的亲水指数和亲水氨基酸分布情况Fig.4 Relationship of hydrophilicity of peptides and ACE activity
疏水性是活性肽的重要分子特性之一,肽段的亲水疏水值可能会影响其与ACE结合的效果。由图4a可见,据本研究涉及的ACE抑制肽统计可知,其肽段整体亲水性值小于0的有465条,占所分析总数的67.29%;整体亲水性值大于0的有181条,占所分析总数的26.19%。说明这些ACE抑制肽中,亲水性较弱而疏水性相对较强的肽段所占比重较大。图4b显示,ACE抑制肽氨基酸组成中所含有的亲水氨基酸含量并不高,肽链中亲水氨基酸百分比含量小于50%的567条肽,在全部统计肽段中占比82.05%;而亲水氨基酸含量大于或等于50%的肽段只有124条,在全部统计肽段中占比17.94%。即大部分的ACE抑制肽中亲水性氨基酸含量较低。
上述这些结果可以作为实际研究中的理论参考,在筛选确定ACE抑制肽过程中明确研究方向,制定制备策略等方面具有良好的指导作用。
2.5 核桃ACE抑制肽的虚拟酶切产物及分析
核桃种子储藏蛋白的分子量为58 144.5 Da,理论等电点为7.0[7-8]。平均亲水性是根据多肽序列中所有氨基酸残基以及它们所带不同修饰基团的亲/疏水值计算的。经过分析,核桃种子储藏蛋白分子的平均亲水性值是0.3,亲水氨基酸残基在分子中所占的比例为49%。从这个蛋白质的平均亲水性值,可以判断其在水相体系中的溶解性比较好。
本研究根据核桃种子储藏蛋白的氨基酸序列,采用 PeptideCutter数据库工具(https://web.expasy.org/peptide_cutter/)[7],进行其虚拟酶切位点分析。虽然中性蛋白酶和碱性蛋白酶也属于内切酶,在目前食品工业中广泛用于酶解蛋白质生产活性肽,但由于这两种酶切位点几乎没有序列特异性,随机性很强,很难控制切割的氨基酸残基位置,产物难以预测,所以在选择虚拟酶切的蛋白酶时不考虑这两种酶。从PeptideCutter数据库中,选取Arg-C蛋白酶[10]、Asp-N端肽酶[11]、Asp-N端肽酶+N端Glu[12]、含半胱氨酸的天冬氨酸蛋白水解酶[13]、高专一性胰凝乳蛋白酶[14]、低专一性胰凝乳蛋白酶[15]、谷氨酰内肽酶[16]、胃蛋白酶(pH1.3)[17]、胃蛋白酶(pH>2)[18]、脯氨酸肽链内切酶[19]、蛋白酶 K[20]、葡萄球菌肽酶I[21]、嗜热菌蛋白酶[22]、胰蛋白酶[23]这14种较为常见的蛋白酶进行虚拟酶切分析。
研究表明,组织蛋白酶C是一种巯基蛋白酶,可以水解氨基端双肽,可通过K、R或P氨基端作为第二或第三个氨基酸进行封闭;胰凝乳蛋白酶是一种丝氨酸蛋白酶,特异性水解位点是在F-Phe、T-Thr或Y-Tyr之后;内肽酶Arg-C是一种丝氨酸蛋白酶,特异性水解位点是在R-Arg之后[10];内肽酶Asp-N是一种金属蛋白酶,特异性水解位点是在D-Asp和C-Cys半胱氨酸之前[11];内肽酶Glu-C是一种丝氨酸蛋白酶,特异性水解位点是在E-Glu/Gln或D-Asp之后;内肽酶Lys-C是一种丝氨酸蛋白酶,特异性水解位点是在K-Lys之后;蛋白酶K是一种丝氨酸蛋白酶,特异性水解位点具有广泛特异性[20];枯草杆菌蛋白酶是一种丝氨酸蛋白酶,水解位点无特异性;胰蛋白酶是一种丝氨酸蛋白酶,特异性水解位点是在K-Lys或R-Arg之后[23]。胃蛋白酶对蛋白质的酶切位点具有一定的氨基酸序列特异性,优先水解N-末端或C-末端为芳香族氨基酸(Phe、Trp和Tyr)或Leu的肽键。关于不同蛋白酶的酶切位点与核桃种子储藏蛋白氨基酸序列之间的对应关系如表1和图5所示。

表1 核桃种子储藏蛋白的理论酶切位点(100%可能性)Table 1 Theoretical enzyme cutting site of walnut seed storage protein(100% possibility)
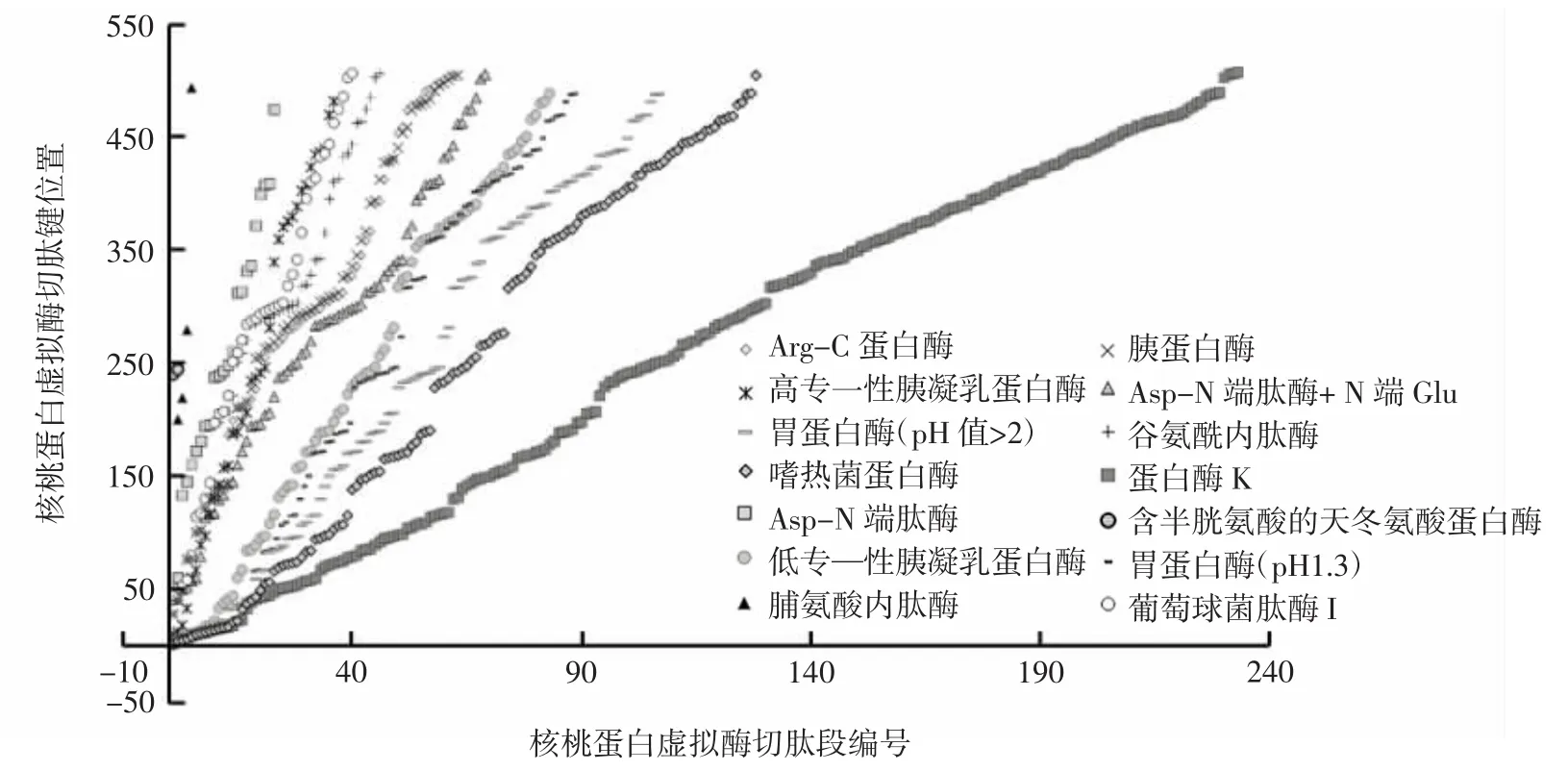
图5 核桃种子储藏蛋白的虚拟酶切位点分析Fig.5 Analysis of the virtual enzyme cutting site of walnut seed storage protein
3 结论
本研究建立了ACE抑制肽分子结构信息与ACE抑制活性之间的内在规律性联系。其中,分子量小于1 000 Da尤其是500 Da以下的ACE抑制肽占据优势地位,并且 Pro、Tyr、Leu、Phe和 Ala这几种氨基酸在ACE抑制肽分子末端出现的频率较高;这些肽分子的理论等电点在pH 5~7区间分布较多,而且这些肽中亲水性氨基酸含量通常较低。另外,核桃储藏蛋白的虚拟酶切分析表明,组织蛋白酶K的酶切位点最多,约占总肽键数量的50%以上;人体消化道中常见的胰蛋白酶、胃蛋白酶、糜蛋白酶,在酶解制备核桃蛋白多肽中也表现出良好的应用前景。该研究为以核桃蛋白为原料的ACE抑制肽的制备提供了一定的理论参考。
