花生主要品质性状的主基因+多基因遗传分析
2021-08-30牟大林韩笑李雪莹姚丹杨松楠曲艺伟梁嘉宁张
牟大林韩 笑李雪莹姚 丹杨松楠曲艺伟梁嘉宁张 君
(吉林农业大学农学院,吉林 长春 130118)
花生主要分布在美洲、非洲和亚洲[1]。我国是世界上最大的花生生产国和出口国,目前我国花生油的市场需求量与实际生产缺口量达到了50万t,随着社会发展和百姓生活水平的提高,研究者提出应该加强高油花生育种及花生品质性状改良[7]。此前研究花生大多归于对种质筛选[2]、性状相关性[3-4]及亲本配合力[5-6]的研究。封海胜[8]、夏虹[9]、姜慧芳[10]等先后利用不同杂交组合分析了花生品质性状的遗传规律,并培育出了高油、高蛋白等优质专用型花生品种。夏友霖等认为不同花生蛋白质和脂肪含量配合力和相对遗传力不同[11]。对花生蛋白质和脂肪遗传研究发现其基因效应以加性效应为主[6,12]。
基于此,本试验采用盖钧镒等[13-14]建立的混合遗传模型,深入探讨花生主要品质性状的遗传规律,分析数量性状的最适遗传模型,明确遗传因素对蛋白质和脂肪的贡献率,为花生品质育种提供理论支撑,同时为探索高脂肪、高蛋白基因的QTL定位和分子辅助标记育种研究奠定基础。
1 材料和方法
1.1 供试材料
以潍花8号(P1)和12L49(P2)作亲本,配置5个家系世代。2017年春天在吉林农业大学的试验地进行杂交,获得F1代花生种子,分成2份,1份进行保留,另1份在冬天进行南繁加代,获得F2代的花生种子;2018年,种植一部分F2代花生种子,另一部分种子保留,获得F2:3代的衍生系;2019年,在学校试验基地进行5个家系世代的材料种植,在潍花8号和12L49亲本中分别随机抽取25份和30份材料,在F1代、F2代群体中各抽取25份和150份材料,从F2:3代群体中,抽取140个家系材料,从每个家系中随机取5株,合计1850份材料。
1.2 蛋白质和脂肪测定
成熟时按家系剔除边际效应进行收获,自然晾晒,风干后进行室内考种并测量数据。利用吉林农业大学农学院谷物分析实验室Buchi公司的近红外光谱仪N500,测定5个世代群体品质性状含量。以5个家系各抽取15粒植株的平均值作为脂肪和蛋白质含量,运用Ri386 3.4.4分析软件进行测量数据的初步分析。
1.3 统计分析方法
利用多个世代联合分析的方法[15-16],对花生5个世代的脂肪含量和蛋白质含量进行数据分析,使用IECM 算法和极大似然值法估算各有关参数,根据适合性测验和AIC准则,以显著水平数最少的为最适模型,同时估计出各相关品质性状的主基因、多基因方差、效应值和遗传参数值。最后根据遗传模型和遗传率公式(1)和(2)进行计算。
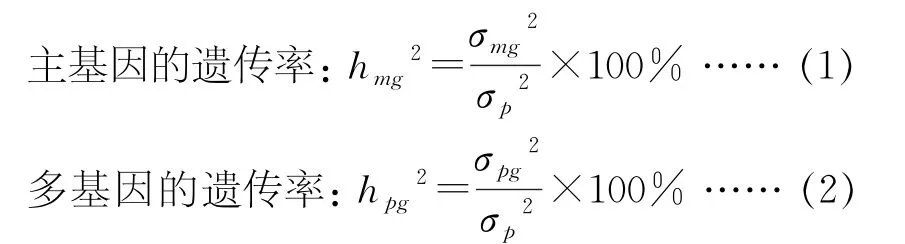
2 结果与分析
2.1 脂肪和蛋白质含量遗传变异
表1中显示花生脂肪和蛋白质品质性状都有一定的变异。潍花8号和12L49平均蛋白质含量分别是23.54%和25.15%,在F2:3代产生了变异和超亲遗传现象,变异幅度在16.51%~27.85%间。亲本中脂肪含量分别为45.87%和50.62%,在F2:3群体中存在着超亲遗传的现象,变异幅度在41.52%~53.96%间,群体平均值在双亲之间,并偏向较高值的亲本。两个性状的偏度分别是0.73和-0.61,并且峰度分别是0.15和0.20,绝对值都比较小,所以群体中脂肪和蛋白质含量属于数量性状分布特征。
2.2 遗传模型
采用群体联合数量性状分离分析方法对脂肪含量和蛋白质含量进行“主基因+多基因”混合遗传模型分析。利用R 软件进行遗传模型分析运算,计算出脂肪和蛋白质的24种遗传模型,得到极大对数似然函数值和 AIC 值,从中选取AIC值较小的三个模型为候选模型(表1)。其中蛋白质含量AIC值较小的3个模型是:MX1-AD-ADI(一对加性—显性主基因+加性—显性—上位性多基因)、MX1-A-AD(一对加性主基因+加性—显性多基因)、MX1-EAD-AD(一对完全显性主基因+加性—显性多基因),AIC值分别为4037.012、4078.602、4080.604,3个模型都属于1对主基因+多基因,说明此组合蛋白质含量的遗传是受一对主基因+多基因控制。

表1 亲本和F2:3 群体脂肪含量和蛋白质含量性状变异Table 1 Variation of fat and protein content in parents and F2:3 population
脂肪含量AIC值较小的3个模型是:PG-AD(加性—显性多基因)、2MG-ADI(2 对加性—显性—上位性主基因)、PG-ADI(加性—显性—上位性多基因),其模型的AIC 值分别为2945.511、2953.324、2955.496,3个模型中有一个显示主基因遗传,另外两个显示多基因遗传控制。
蛋白质3个模型中,MX1-A-AD有13个统计量达到显著水平,MX1-EAD-AD、MX1-AD-ADI有12个统计量达到显著水平,对两个模型进行极大似然比测验(LRT),分析结果模型间有显著差异而且MX1-AD-ADI模型AIC值最小,因此MX1-ADADI为最适模型。所以,确定蛋白质性状的遗传模型为MX1-AD-ADI模型,即1对加性—显性主基因+加性—显性—上位性多基因遗传。
分析表3不同遗传模型AIC值显示,脂肪AIC最小的3个模型中,PG-AD、PG-ADI和2MG-ADI模型均有8个统计量达到显著水平,对三个模型进行极大似然比测验(LRT),结果显示模型有显著差异,而且PG-AD模型AIC值最小,因此PG-AD为脂肪的最适遗传模型,即多基因加性—显性遗传。

表3 花生脂肪和蛋白质含量不同遗传模型的AIC值Table 3 AIC values of different genetic models for fat content and protein content of peanut
2.3 遗传参数估计
表4可见,花生蛋白质的群体均值m=21.99,主基因表现出负向加性效应,多基因的显性效应为[h]=0.6563,而加性效应值是[d]=-1.2126,显性效应明显大于加性效应,在F2代和F2:3代中主基因的遗传率分别为27.0234%和30.2766%;多基因遗传率分别是37.7856%和38.1190%,多基因遗传率76.9046%比主基因遗传率57.3000%高出19.6046%。

表4 蛋白质和脂肪含量遗传参数估计Table 4 Genetic parameter estimation on protein content and fat content of peanut
脂肪含量遗传估计的群体均值m=49.4827,多基因的加性效应[d]=1.1248,F2和F2:3代中无主基因遗传率,而多基因的遗传率分别是32.0843%和32.6325%,所以花生脂肪遗传主要受到微效多基因的控制。
3 讨论与结论
Elston等[17]和Stewart等[18]曾经提出一个主基因+多基因遗传模型,并且在遗传分析中得到了大量应用。王建康[15]拓展了多个世代分离的联合分析方法,该方法适用不同作物脂肪和蛋白质等性状的遗传分析。陈四龙[19]等分析花生脂肪含量的主基因+多基因遗传效应,得出不同的亲本杂交组合在遗传中存在着差异的结论,既存在着主基因+多基因遗传效应,也存在着多基因加性遗传效应,部分结论与本研究相同。
本试验研究中得出蛋白质含量受到一对“加性—显性主基因+加性—显性—上位性”多基因遗传,主基因存在负的加性效应,加性效应为负值(d=-0.16)。多基因加性效应值[d]=-1.21,显性效应值为0.6563,加性效应大于显性效应。在F2和F2:3代,主基因遗传率分别为27.02%、30.27%,遗传力较低,多基因遗传率分别为37.78%、38.11%,主基因遗传率和多基因遗传率分别为57.29%、75.89%,所以在晚世代进行选择时,选择效率较高,采用杂交育种方法改良蛋白质含量的潜力较大。脂肪含量为多基因遗传,在F2和F2:3代中,多基因的遗传率分别为32.0843%和32.6325%,F2和F2:3家系群体主基因遗传率较低,因此选择适宜的育种材料可提高脂肪含量。
花生蛋白质和脂肪含量性状受多基因遗传控制,说明进行多世代回交有利于品质育种。本试验虽得到一些有意义的结论,但复杂的遗传机制并非本试验结果所能完全解释,今后还需结合更加科学的设计方案进行更深层次探讨,以期获得更完善的遗传效应规律。
