叶面施锌对马铃薯叶片光合特性、超微结构及产量的影响
2021-08-26索海翠王丽李成晨刘计涛安康李小波
索海翠 王丽 李成晨 刘计涛 安康 李小波


摘 要:為明确锌对马铃薯叶片光合特性及超微结构的影响,以马铃薯品种‘粤引85-38为试验材料,在出苗后第40 天,叶面喷施5个不同浓度ZnSO47H2O溶液[0(T0)、3.5 mmol/L(T1)、7.0 mmol/L(T2)、14.0 mmol/L(T3)和28.0 mmol/L(T4)]。结果显示,与缺锌处理T0相比较,较低浓度T1和T2处理显著提高了马铃薯叶片的SPAD值和净光合速率(Pn),显著增加了地上部鲜重、地上部干重、薯块鲜重及叶片和薯块锌含量;但当处理浓度达到T4时,马铃薯叶片SPAD值和Pn显著降低,各指标分别与T0处理差异不显著,而最大荧光产量Fm,PSⅡ最大光合量子产量Fv/Fm,非光化学淬灭系数qP和化学淬灭系数NPQ则显著低于T0处理,但叶片和薯块锌含量则显著升高。进一步对叶片显微和超微结构研究显示,T2处理下马铃薯叶片栅栏组织和海绵组织排列整齐,叶绿体形态结构完好,而T0和T4处理降低了叶片表皮厚度,增加了细胞间隙。T0处理下淀粉粒增多,叶绿体膨胀短缩,基粒片层模糊不清;T4处理下细胞凹陷,叶绿体形状改变。以上结果表明,在本研究中,T1~T2处理为较为适宜马铃薯生长的锌浓度。本研究从生理上揭示了锌缺失和过量对马铃薯光合作用和生长发育的影响,为马铃薯田间叶面喷施锌肥提供一定的理论参考。
关键词:马铃薯;叶面施锌;光合作用;超微结构
中图分类号:S532 文献标识码:A
Effects of Leaf-Spraying Zinc Treatment on Photosynthetic Characteristics, Ultrastructure of Potato Leaves and Yield
SUO Haicui, WANG Li, LI Chengchen, LIU Jitao, AN Kang, LI Xiaobo*
Crops Research Institute, Guangdong Academy of Agricultural Sciences / Guangdong Province Key Laboratory of Crop Genetic Improvement, Guangzhou, Guangdong 510640, China
Abstract: In order to clarify the effects of zinc on the photosynthetic characteristics and ultrastructure of potato leaves, potato variety ‘Yueyin 85-38 was used. Five ZnSO47H2O solutions with different concentrations [0 (T0), 3.5 mmol/L(T1), 7.0 mmol/L(T2), 14.0 mmol/L(T3) and 28.0 mmol/L(T4)] were sprayed on the leaves at the 40th day after seedling emergence. Compared with T0 treatment, low concentration treatments T1 and T2 significantly increased the leaf SPAD value and net photosynthetic rate (Pn), and the aboveground fresh and dry weight, fresh tuber weight as well as leaf and tuber zinc content were also significantly increased. When the spraying concentration reached T4, however, the potato leaf SPAD and Pn were significantly reduced, which was not significantly different with that of T0, and the maximum fluorescence yield Fm, PS Ⅱ maximum photosynthetic quantum yield(Fv/Fm), non photochemical quenching coefficient (qP) and chemical quenching coefficient (NPQ) were significantly lower than that of T0, while the leaf and tuber zinc content were increased significantly. Leaf microstructure and ultrastructure showed that under T2 treatment, palisade tissue and sponge tissue of potato leaves were neatly arranged, chloroplast morphology and structure were intact. While under T0 and T4 treatments, the uniformity of palisade tissue was decreased, and intercellular was increased. Under T0 treatment, starch granules increased, the chloroplast expanded and shrank, the granule lamella was obscure. Under T4 treatment, cell and chloroplast membrane structure was damaged. The zinc concentration of T1 and T2 was suitable for potato growth. The study revealed the physiological effects of zinc deficiency and excess on the photosynthesis of potato, and could provide theoretical reference for the application of zinc fertilizer on potato in the field.
Keywords: potato (solanum tuberosum L.); leaf-spraying zinc; photosynthesis; ultrastructure
DOI: 10.3969/j.issn.1000-2561.2021.07.020
锌是植物体所必需的微量营养元素之一,作为六大类功能酶中的辅助因子成分,能够调节酶的活性,广泛参与植物体多种生长发育代谢过程,包括蛋白质、核酸、碳水化合物和磷脂的合成与代谢、植物生长激素的调控、应对逆境胁迫以及光合作用等[1-3]。锌对光合作用的影响主要有以下几个方面:首先,锌作为叶绿体的组成成分,对于光合色素的合成、维持叶绿体结构和叶绿体膜系统的完整性具有重要作用[4],缺锌或锌过量使玉米叶片叶绿素含量下降、叶绿体数量减少和结构发育受阻[5];番茄叶绿体膜系统损伤,MDA水平增加[6]。其次,缺锌或锌胁迫可导致光合特性和荧光特性的改变,缺锌会造成苹果树叶片净光合速率(Pn)、气孔导度(Gs)下降,胞间CO2浓度(Ci)升高[7],缺锌使玉米叶片Pn、Tr、Gs和PSⅡ最大光合量子产量(Fv/Fm)下降[4-5]。在桑科和十字花科研究中表明,缺锌降低了叶绿素SPAD值,降低了Pn、Gs,抑制了电子传递(ETR),升高了初始荧光Fo[8]。在高浓度锌胁迫下,黑麦叶片的Fv/Fm和PSⅡ的实际光合量子产量ΦPSⅡ下降[9]。据报道,缺锌引起的光合速率下降可能是Gs降低或碳酸酐酶活性下降引起的[4,10]。再次,锌对叶片显微结构及超微结构也有影响,如缺锌玉米叶肉细胞膜皱缩,叶绿体数目减少,淀粉粒增多,叶绿体收缩膨胀变形,外膜消失[4,11]。研究表明,随缺锌程度加重,苹果叶片厚度增加,叶肉细胞排列松散,叶绿体缩短膨胀,基粒片层排列杂乱甚至模糊不清,淀粉颗粒小且数目多[7,12]。
马铃薯(Solanum tuberosum L.)粮菜兼用,营养全面,是世界第四大粮食作物。锌是马铃薯生长必不可少的微量元素之一,施锌能提高马铃薯产量和干物质含量[13-14],同时能提高块茎还原性糖、维生素C及锌含量等[15-16]。但关于锌对于马铃薯的光合作用,尤其是叶片光合特性和超微结构方面的影响却鲜有报道。本试验以马铃薯品种‘粤引85-38为材料,进行不同浓度锌溶液喷施处理,通过研究锌对马铃薯叶片荧光特性、超微结构、块茎产量及锌含量等指标的影响,探究锌对光合作用影响的生理机制,为马铃薯田间安全喷施锌肥提供理论基础。
1 材料与方法
1.1 材料
试验在广东省广州市白云区广东省农业科学院试验基地网棚中进行。供试马铃薯品种为广东省主栽品种‘粤引85-38,脱毒种薯由广东省农业科学院作物研究所提供。试验采用砂培盆栽培养,分别选择直径为1~2 mm(12.14 kg/盆)和0.4~0.7 mm(4.27 kg/盆)的石英砂颗粒(购自广东半岛石英砂厂)按3∶1比例混合均匀后装入直径25 cm,有效容积约为11 L的花盆。经0.5%的高锰酸钾消毒处理后的切刀将种薯切成重约25 g的薯块,每个薯块带1~2个芽眼。随后用混匀的50%多菌灵可湿性粉剂与滑石粉(比例为1∶20)进行拌种,拌种晾干后的薯块放入盆中,每盆一个薯块,覆盖石英砂厚度约2 cm,去离子水浇透。
出苗后,每3~4 d用不含ZnSO4的Hoagland培养液浇灌幼苗。出苗后40 d分别叶面均匀喷施4个浓度的ZnSO4·7H2O溶液[3.5 mmol/L(T1)、7.0 mmol/L(T2)、14.0 mmol/L(T3)和28.0 mmol/L(T4)],以喷施去离子水作为缺锌处理(T0)。试验处理采用随机区组试验设计,3次重复。处理后10 d进行叶绿素SPAD值、光合参数、荧光参数等的测定。生长90 d收获马铃薯地上部和薯块,分别测定其干鲜重,同时测定叶片和薯块中的锌含量。
1.2 方法
1.2.1 叶片和薯块锌含量测定 马铃薯植株收获后分为叶片和薯块2部分,用去离子水冲洗干净,于烘箱105 ℃杀青30 min,然后75 ℃烘干至恒重。将烘干样品粉碎磨细,植物叶片和薯块的锌含量测定采用火焰原子吸收分光光度法[17],测定仪器为ZA3300型原子吸收分光光度计,所选参数波长为213.8 nm(日本HITACHI公司)。
1.2.2 SPAD值测定 采用SPAD-520型手持便携式叶绿素测定仪(日本Monlta公司)测定。选取各植株顶端完全展开、成熟的第二片葉,在叶片主脉两侧从叶尖到叶基部均等测6个点,取其平均值,测定前用标准色板校正仪器基准数值。
1.2.3 光合参数测定 采用CI-340手持式光合作用测定系统(美国CID公司),于晴朗上午10:00– 12:00,选取不同处理中马铃薯植株第3片叶测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。测定光强为1300 μmol/(m2·s)、CO2浓度为400 μL/L,叶片温度在25 ℃~30 ℃之间。
1.2.4 叶绿素荧光动力学参数测定 采用PAM-2500型便携式调制叶绿素荧光仪(德国WALZ公司)对不同浓度处理的叶片测定光适应下的最小荧光产量(Fo?)、稳态荧光产量(Fs)及光下最大荧光产量(Fm?);测定是在整株植株上进行,使用叶夹DLC-8夹住叶片,关闭滑片,暗适应30 min,测定叶片暗适应下的初始荧光产量(Fo)及最大荧光产量(Fm)。PSⅡ的最大光合量子产量Fv/Fm=(Fm–Fo)/Fm。打开滑片,测定光适应下PSⅡ的实际光合量子产量ΦPSⅡ=(Fm?–Fs)/Fm?;光化学淬灭系数qP=(Fm?–Fs)/(Fm?–Fo?);非光化学淬灭系数NPQ=(Fm–Fm?)/Fm?。
1.2.5 马铃薯叶片显微和超微切片制作 (1)半薄切片制作。把不同浓度锌处理的马铃薯叶片分别切成1 mm ×3 mm的小块,采用戌二醛和锇酸双固定法制备电镜样品,采用徕卡超薄切片机切片(德国徕卡,EMUC7),切片厚度是800~1000 nm,切片用1%苯胺蓝染色晾干后,在奥林巴斯光学显微镜(日本Olympus)下使用目镜测微尺观测上下表皮厚度、栅栏组织和海绵组织厚度。每个结构参数均为25视野范围内测定平均值。其中,栅栏组织和海绵组织的厚度比(栅/海)=栅栏组织厚度/海绵组织厚度。(2)超薄切片制作。制片方法同上,切片厚度为50~70 nm,切片经醋酸双氧铀和柠檬酸铅溶液复染干燥后上HITACHI透射电子显微镜(日本HITACHI,HT7700)观察。
1.3 数据处理
试验数据均采用Microsoft Excel 2016軟件进行平均值和标准误的计算;采用DPS数据处理软件进行单因素方差分析,数据的差异显著性检验釆用Duncans方法进行多重比较。
2 结果与分析
2.1 不同锌溶液喷施处理对马铃薯叶片叶绿素SPAD值的影响
由图1可知,随着处理锌浓度的增加,马铃薯叶片叶绿素SPAD值呈现先升高后降低的变化规律。与缺锌处理T0相比较,T1和T2处理显著提高了叶片叶绿素SPAD值,分别提高了4.97%和6.00%,而T3和T4处理与T0处理之间差异不显著。但与T2处理相比较,T3和T4处理则显著降低了叶片叶绿素SPAD值,分别降低了3.31%和4.78%。
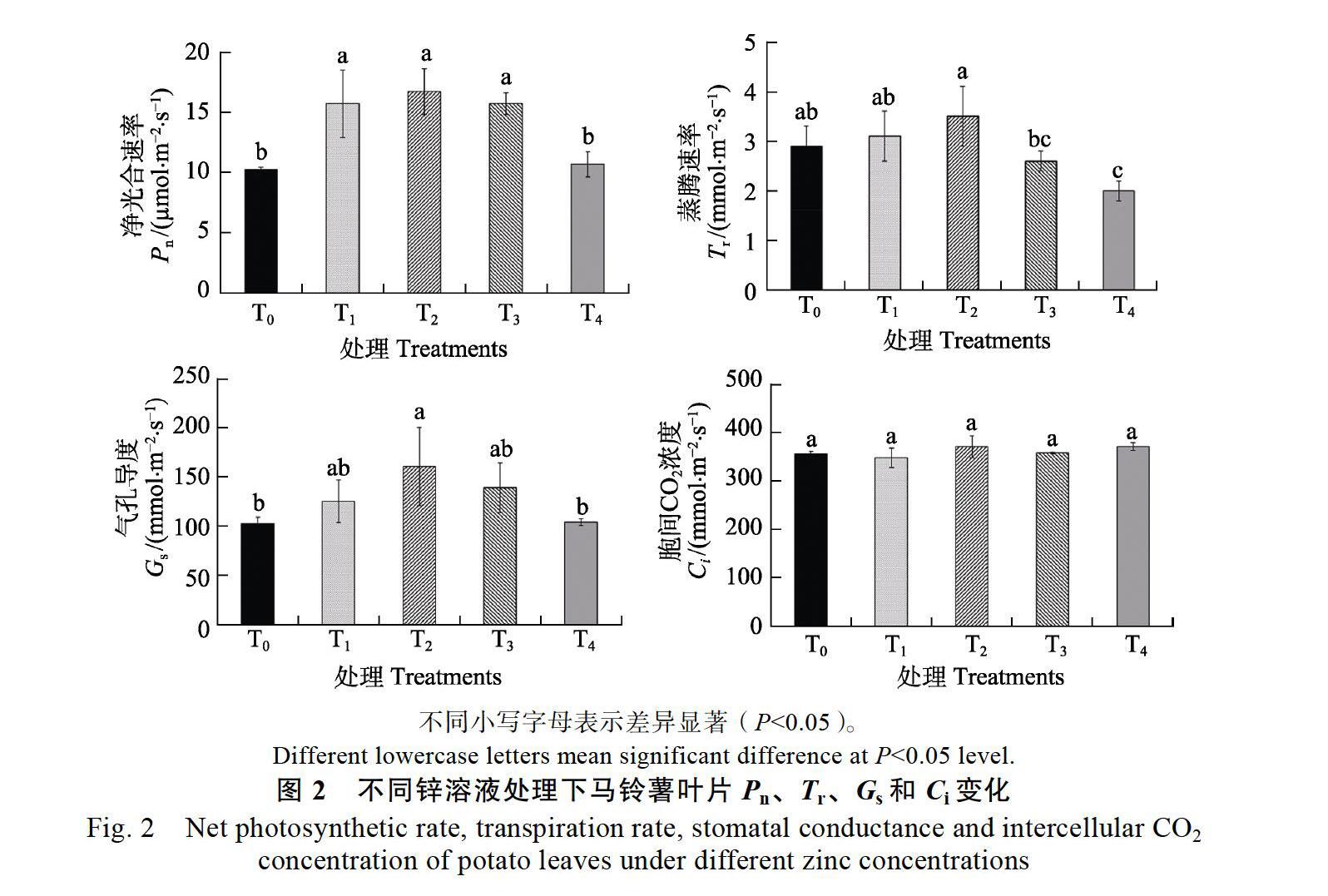
2.2 不同锌溶液处理对马铃薯叶片气体交换参数的影响
由图2可知,随着处理锌浓度的增加,马铃薯叶片Pn呈现先升高后降低的趋势,与T0处理相比较,T1、T2和T3处理显著提高了马铃薯叶片的Pn,分别比T0增加了46.98%、56.28%和46.98%,但T4与T0处理之间叶片Pn差异不显著;马铃薯叶片Tr也呈现先升高后降低的趋势,缺锌处理T0与低浓度T1、T2和T3之间差异不显著,但与T0处理相比较,T4处理显著降低了Tr,降低了45%;随着处理锌浓度的增加,马铃薯叶片的Gs与Pn、Tr相似,呈现出先升高后降的趋势,与T0处理相比较,T2处理显著提高了Tr,提高了55.83%,而T1、T3、T4与 T0间的Gs差异不显著;不同的是,锌浓度处理对马铃薯叶片Ci影响不大,不同锌浓度处理间Ci的差异均不显著。
2.3 不同锌溶液处理对马铃薯叶片荧光参数的影响
由表1可知,分别与缺锌处理T0的叶片不同荧光参数相比较,T1、T2和T3处理叶片的Fo显著降低,分别降低了10.53%、5.26% 和15.79%;T3和T4叶片Fm显著降低,分别降低了19.10%和10.82%;T4处理叶片的Fv/Fm显著降低,降低了6.67%;T3和T4处理下qP显著降低,均降低了10.71%;在NPQ上,与缺锌处理T0相比较,除T2处理显著提高了叶片NPQ,提高了25%外,其他各处理与T0之间在叶片NPQ上差异不显著。
2.4 不同锌处理对马铃薯干鲜重的影响
不同浓度锌溶液处理对马铃薯地上部鲜重、干重和薯块鲜重的影响如图3所示。与缺锌处理T0相比,处理T1、T2和T4显著提高了地上部鲜重,分别提高了147.23%、81.06%和101.4%,其中T1处理下地上部鲜重最高,达到83.02 g,显著高于处理T2,但与处理T4差异不显著,处理T3和T0之间地上部鲜重差异不显著;此外,与缺锌处理T0相比,T1处理显著提高了马铃薯地上部干重,提高了100.81%,而其他各处理与T0处理间差异均不显著。在薯块鲜重上,缺锌处理T0薯块鲜重最低,与缺锌处理T0相比,各施锌处理均显著提高了薯块鲜重,T1、T2、T3和 T4处理分别比T0处理增加了54.03%、110.26%、107.83%%和72.28%,其中,T2、T3处理下薯块鲜重最高,分别为74.56 g和73.7 g,但二者间差异不显著,与T2处理相比,T1和T4处理下薯块鲜重有所降低,但T4与T2处理差异不显著,而T1与T2处理差异显著。
2.5 不同锌处理对马铃薯叶片和块茎锌含量的影响
随着喷施锌浓度增加,叶片和块茎的锌含量呈逐渐增加趋势,缺锌处理T0的叶片和块茎的锌含量最低,其他各施锌处理均显著提高了叶片和块茎的锌含量,其中,T4处理下叶片和块茎的锌含量均达到最大值,分别为1144 mg/kg和54.3 mg/kg,显著高于其他各处理(图4)。对于叶片锌含量,处理T2和T3锌含量次之,二者之间差异不显著,而T1处理叶片锌含量显著低于T2和T3,T0最低。而对于薯块锌含量,不同处理间锌含量表现为:T4>T3>T2>T1>T0,且各处理间差异均达到显著水平(图4)。
2.6 不同锌处理对马铃薯叶片显微和超微结构的影响
综合以上结果,可以看出,在5个不同锌浓度处理中,处理T1和T2呈现出较高的叶绿素SPAD值、Pn以及产量表现。在本研究中,T1和T2浓度范围(3.5~7.0 mmol/L)为马铃薯生长较为适宜的锌浓度。因此,以T2处理作为对照,T0和T4分别为缺锌和过量锌处理,进一步研究锌对马铃薯叶片显微和超微结构的影响。
从表2和图5可以看出,与T2处理相比,缺锌处理T0植株叶片上表皮厚度显著变薄,表皮厚度由T2处理的1.63 μm减少到1.35 μm,但下表皮厚度没有显著差异。同样,与T2处理相比,缺锌处理T0栅栏组织变薄,厚度显著减少了30.56 μm,栅/海比显著降低了0.46,但海绵组织厚度却显著增加了8.93 μm,。与T2处理相比,锌过量T4处理表皮细胞排列更为紧凑,上、下表皮厚度与海绵组织厚度差异不显著,但栅栏组织厚度和栅/海比显著降低。
细胞超微结构结果显示,与T2处理相比,缺锌T0处理马铃薯叶片细胞内淀粉粒变小且增多,叶绿体体积变小、短缩,成近圆形,排列紧密,基粒片层排列杂乱且模糊不清;而过量锌T4处理下,细胞凹陷,细胞膜、液泡膜结构损坏,叶绿体游离于细胞液中,叶绿体膜结构不清晰,叶绿体形状不规则,基粒片层松散(图5)。
3 讨论
在实际生产中,施用锌肥能够提高作物的产量和品质,如小麦中,进行叶面喷施锌肥提高了小麦产量和籽粒锌含量[18-19]。马振勇等[15]在马铃薯生育期进行2次喷施锌肥后,叶片中锌浓度比不喷锌处理分别高27.94和43.89 mg/kg,收获后块茎中的锌浓度比不喷锌高28.70%。本研究中,与不施锌肥(T0)相比较,不同浓度施锌处理能够显著提高马铃薯叶片和块茎中的锌浓度,在T1处理下,叶片和薯块锌浓度比缺锌T0处理分别高321.19%和33.32%。此外,与T0不施锌肥相比较,随着锌处理浓度增加,马铃薯产量逐渐增加,在T2处理下达到最大值,但当锌浓度高于T4处理(锌浓度≥28mmol/L)时,马铃薯薯块鲜重有所降低。前人研究表明,合理施用锌肥可以提高马铃薯块茎产量[13-14],而过量施锌则导致马铃薯减产[20-21],与本研究结果一致。
前人研究锌对植物光合作用的影响,多为单独缺锌或者锌过量的影响[22-23]。本研究设置了一系列锌浓度处理,从光合参数,荧光参数以及叶片显微和细胞超微结构方面研究了从缺锌到过量锌处理对马铃薯光合作用的影响,是对前人研究的扩展和补充。研究结果显示,在5个不同锌浓度处理中,处理T1和T2呈现出较好的叶绿素SPAD值、气体交换参数、荧光参数以及产量表现。在本研究中,确定处理T1到T2浓度区间(3.5~7.0 mmol/L)为马铃薯生长较为适宜的锌浓度。与较为适宜处理浓度T2相比,缺锌处理T0和高锌处理T4下,马铃薯叶片叶绿素SPAD、Pn和Gs显著下降,而Ci在各处理间差异不显著,据报道,引起Pn下降的因素主要有2类,一类是气孔因素,主要是受气孔数量、气孔孔径和气孔开度等影响,通过降低Ci而减少光合作用的碳源[10,24] ;另一类为非气孔因素,主要是受内部酶活性和光合组分控制[25-26]。在本研究中,不同浓度锌处理对马铃薯叶片Ci影响不显著,因此,缺锌和高锌处理引起的马铃薯叶片Pn下降为非气孔因素,与前人在谷子[27]、辣椒[28]中的研究一致[27-29]。
叶绿体是植物细胞内进行光合作用的细胞器,叶绿素荧光参数在研究植物的光化学效率、光抑制与光破坏防御等方面具有重要作用。本研究结果表明,与T2处理相比较,缺锌T0处理显著降低了马铃薯叶片叶绿素SPAD值、光合速率Pn、气孔导度Gs和非光化学淬灭系数NPQ,显著减少了地上部鲜重和薯块鲜重。前人对苹果的研究表明,随缺锌程度加重,叶片叶绿素SPAD值、光合速率显著下降,Fo上升,同时Fv/Fo、ΦPSⅡ、qP及Fv/Fm显著下降[7,12]。此外,缺锌对植物叶片的显微结构也具有一定的影响。研究表明,缺锌导致了苹果叶片厚度增加,栅栏组织细胞变宽,海绵组织细胞间空隙的增加[7,12];隨着缺锌程度加重,山核桃叶片上下表皮厚度增加,栅栏组织、海绵组织厚度减少[30]。本研究结果显示,缺锌降低了马铃薯叶片上表皮厚度,降低栅栏组织厚度,海绵组织细胞间隙变大。可见缺锌对不同物种的叶片上下表皮厚度影响不同,暗示不同物种叶片对缺锌胁迫的响应可能存在一定差异。此外,本研究结果也显示了缺锌对马铃薯叶片超微结构的影响,缺锌下叶片中的叶绿体缩短膨胀,排列紧密,基粒片层排列杂乱甚至模糊不清,淀粉颗粒小且数目多,与前人在玉米[4],苹果[12]中研究结果基本一致。表明缺锌致使叶绿体发育不良,导致叶绿体结构和功能异常。
当锌元素大量存在植物的生长环境中,植物器官中锌浓度超出植物对于锌需求的最佳浓度,从而可能导致植物的毒性作用和代谢紊乱[31]。如在番茄幼苗中,高浓度锌胁迫(≥50 ?mol/L)处理导致番茄幼苗Fm、Fv/Fm、Fv/Fo均有不同程度下降[22]。高锌胁迫下,汉麻的Fv/Fm、ΦPSⅡ、qP和ETR随着锌浓度的升高而呈下降趋势[32]。本研究中,高锌浓度处理(≥14 mmol/L–1)显著降低了叶片SPAD值、Pn、Gs、Tr,同时也显著降低了Fm、Fv/Fm、qP和NPQ。表明在不同物种中,高锌胁迫都抑制了PSⅡ的光能捕获和转化效率。进一步对细胞显微和超微结构研究表明,高锌胁迫下细胞凹陷,液泡破裂,叶绿体形状不规则,尤其是膜系统包括细胞膜、液泡膜、叶绿体膜和类囊体膜受到损坏。前人研究表明,锌毒害会导致过氧化反应,破坏光合作用的电子传递过程,产生毒害的活性氧类物质(ROS),如:O2–和H2O2,增加了脂质过氧化,从而破坏了膜透性[33]。如在番茄幼苗研究中发现,过量锌胁迫下番茄幼苗叶片的丙二醛(MDA)含量增加[22]。此外,过量锌胁迫下,抗氧化系统即超氧化物歧化酶、抗坏血酸过氧化物酶和谷胱甘肽还原酶被激活[34],但本研究中抗氧化系统酶类是否被激活还有待进一步研究。研究证实缺锌也会使植株产生活性氧类物质(ROS),导致内囊体膜结构损害[4]。但在玉米中研究发现,活性氧类物质的产生与品种锌效率相关,锌效率高的品种在缺锌下过氧化物酶(POD)和超氧化物岐化酶(SOD)活性高于锌效率低的品种,而POD和SOD对活性氧类物质导致的光氧化伤害具有保护作用,在一定程度上降低了高浓度锌对细胞的毒害[23]。因此,推测这可能是本研究中缺锌条件下马铃薯细胞膜结构相对完整的可能原因之一,需进一步研究证实。综上所述,在本研究中,锌过量与锌缺乏对光合作用造成的伤害的机理既有相同又略有不同,相同点在于均导致细胞间隙变大,栅/海比降低,叶绿体性状改变,基粒片层松散等,而不同在于缺锌主要是导致了叶绿体发育不良,致使叶绿体结构和功能异常,而锌过量对马铃薯光合作用的抑制主要是由于锌过量引起的氧化反应,导致细胞膜、液泡膜、叶绿体膜和类囊体膜等膜结构的破坏。最终,缺锌和过量施锌都抑制了马铃薯产量,但与缺锌处理比较,锌过量处理下的马铃薯具有较高的地上部和块茎鲜重,表明马铃薯对于锌毒害具有较强的耐性。
4 結论
适宜的锌浓度(3.5~7.0 mmol/L)能提高马铃薯叶片光合效率,增加马铃薯产量,而缺锌和过量锌(≥14 mmol/L)都抑制了马铃薯叶片的光合效率,而2种处理对光合作用造成的伤害的机理既有相同又略有不同,缺锌下马铃薯叶绿体发育不良,导致叶绿体结构和功能异常,而过量锌胁迫下马铃薯细胞膜和叶绿体膜等膜结构受到破坏。最终,缺锌和过量施锌都影响了马铃薯产量,但与缺锌相比较,锌过量处理对马铃薯产量的影响相对较小。本研究结果可对生产上马铃薯叶面喷施液体锌肥提供一定的理论参考。但由于本研究只分析了锌处理对马铃薯叶片光合生理变化、叶片显微和细胞亚显微结构的影响,对于光合作用中光合酶的活性,及过氧化酶系统中酶的活性等还有待进一步研究。
参考文献
[1] Keram K S, Sharma B L, Sharma G D, et al. Impact of zinc application on its translocation into various plant parts of wheat and its effect on chemical composition and quality of grain[J]. Scientific Research and Essays, 2013, 8(45): 2218-2226.
[2] Abdoli M, Esfandiari E, Alilloo A, et al. Study of genetic diversity in different wheat species with various genomes based on morphological characteristics and zinc use efficiency under two zinc deficient growing conditions[J]. Acta Agriculturae Slovenica, 2019(113): 147-161.
[3] Broadley M R, White P J, Hammond J P, et al. Zinc in plants[J]. New Phytologist, 2007, 173(4): 677-702.
[4] Zhang J Y, Wang S F, Song S H, et al. Transcriptomic and proteomic analyses reveal new insight into chlorophyll synthesis and chloroplast structure of maize leaves under zinc deficiency stress[J]. Journal of Proteomics, 2019, 199: 123-134.
[5] Edson M M, Hugo A R, Julio C L N , et al. Zinc deficiency affects physiological and anatomical characteristics in maize leaves[J]. Journal of Plant Physiology, 2015(183): 138-143.
[6] Cherif J, Derbel N, Nakkach M, et al. Analysis of in vivo chlorophyll fluorescence spectra to monitor physiological state of tomato plants growing under zinc stress[J]. Journal of Photochemistry and Photobiology B: Biology, 2010, 101(3): 332-339.
[7] Fu C X, Li M, Zhang Y, et al. Morphology, photosynthesis, and internal structure alterations in field apple leaves under hidden and acute zinc deficiency[J]. Scientia Horticulturae, 2015(193): 47-54.
[8] Zhao K, Wu Y Y. Effects of Zn deficiency and bicarbonate on the growth and photosynthetic characteristics of four plant species[J]. PLoS One, 2017, 12(1): e0169812.
[9] Bonnet M, Camares O, Veisseire P. Effect of zinc and influence of Acremonium lolii on growth parameters, chlorophyll a fluorescence and antioxidant enzyme activities of ryegrass (Lolium perenne L. cv Apollo)[J]. Journal of Experimental Botany, 2000, 51(346): 945-953.
[10] Sharma P N, Tripathi A, Bisht S S. Zinc requirement for stomatal opening in cauliflower[J]. Plant Physiology, 1995, 107(3): 751-756.
[11] 王盛锋, 高丽丽, 刘自飞, 等. 不同土壤水分供应下锌对玉米叶片超微结构的影响[J]. 中国生态农业学报, 2013, 21(8): 959-965.
[12] 付春霞. 锌对苹果叶片结构发育, 光合及荧光特性的影响[D]. 泰安: 山东农业大学, 2014.
[13] 王延明. 锌肥用量及施用方法对马铃薯产量形成及营养品质的影响[D]. 兰州: 甘肃农业大学, 2014.
[14] 侯叔音. 不同锌肥对旱作马铃薯产量形成及锌素吸收和积累的影响[D]. 兰州: 甘肃农业大学, 2013.
[15] 马振勇, 杜虎林, 刘荣国, 等. 施锌肥对马铃薯干物质积累生理特性及块茎营养品质的影响[J]. 干旱区资源与环境, 2017, 31(1): 148-153.
[16] 李 凯, 张国辉, 郭志乾, 等. 叶面喷施铁锌锰微肥对马铃薯生长品质与产量的影响[J]. 作物研究, 2018, 32(1): 28-34.
[17] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000: 279-281.
[18] Shariatipour N, Seyed-Siamak A, Mohammad M V, et al. Foliar applied zinc increases yield, zinc concentration, and germination in wheat genotypes[J]. Agronomy Journal, 2019, 112 (2): 961-974.
[19] Li M, Wang S X, Tian X H, et al. Improving nutritional quality of wheat grain through foliar Zinc combined with macronutrients[J]. Agronomy Journal, 2018, 110(1): 38-46.
[20] 吉 勇, 乐 春, 陈庭珠, 等. 不同浓度锌处理对马铃薯性状的影响[J]. 现代农业科技, 2016(5): 89-90, 92.
[21] 刘新稳, 孙亮庆, 张丽娟, 等. 不同施锌量对马铃薯植株锌的吸收、积累及薯块产量的影响[J]. 江西农业学报, 2018, 30(6): 35-38.
[22] 孙 卓, 李亚迪, 纪建伟. 番茄幼苗叶绿素荧光参数对锌胁迫的响应[J]. 北方园艺, 2016(24): 41-44.
[23] Khatun M A, Hossain M M, Bari M A, et al. Zinc defi-ciency tolerance in maize is associated with the up-regulation of Zn transporter genes and antioxidant activities[J]. Plant Biology, 2018, 20 (4): 765-770.
[24] 井大炜. 杨树苗叶片光合特性和抗氧化酶对干旱胁迫的响应[J]. 核农学报, 2014, 28(3): 532- 539.
[25] 吴炫柯, 李永健, 李楊瑞. 不同木薯品种气体交换特性及光合酶活性的变化[J]. 亚热带农业研究, 2006, 2(3): 179-183.
[26] Rehman H, Aziz T, Farooq M, et al. Zinc nutrition in rice production systems: a review[J]. Plant Soil, 2012, 361 (1-2): 203-226.
[27] 张伟莉, 张丽光, 杨慧杰, 等. 阔世玛对谷子幼苗叶片光合特性及可溶性物质含量的影响[J]. 核农学报, 2020, 34(6): 1294-1301.
[28] 杨万基, 蒋欣梅, 高 欢, 等. 28-高芸苔素内酯对低温弱光胁迫辣椒幼苗光合和荧光特性的影响[J]. 南方农业学报, 2018, 49(4): 741-747.
[29] Chen W, Yang X, He Z, et al. Differential changes in photosynthetic capacity, 77K chlorophyll fluorescence and chloroplast ultrastructure between Zn-efficient and Zn-inefficient rice genotypes (Oryza sativa) under low zinc stress[J]. Physiologia Plantarum, 2007, 132(1): 89-101.
[30] Kim T, Wetzstein H Y. Cytological and ultrastructural evaluations of zinc deficiency in leaves[J]. Journal of the American Society for Horticultural Science American Society for Horticultural Science, 2003, 128(2): 171-175.
[31] 李 娟, 陳杰忠, 黄永敬, 等. Zn营养在果树生理代谢中的作用研究进展[J]. 果树学报, 2011, 28(4): 668-673.
[32] 保琦蓓, 唐 寅, 田光明. 锌胁迫对汉麻光合特性及叶绿素荧光参数的影响[J]. 安徽农业科学, 2016, 44(28): 85-88, 108.
[33] Dietz K J, Ismail T, Anja K L. Redox- and reactive oxygen species-dependent signaling into and out of the photosynthesizing chloroplast[J]. Plant Physiology, 2016, 171(3): 1541-1550.
[34] Jain R, Srivastava S, Solomon S, et al. Impact of excess zinc on growth parameters, cell division, nutrient accumula-tion, photosynthetic pigments and oxidative stress of sugarcane (Saccharum spp.)[J]. Acta Physiologiae Plantarum, 2010, 32(5): 979-986.
责任编辑:白 净
