巴西橡胶树‘RY7-33-97’不同级别死皮树皮结构分析
2021-08-26卢亚莉张世鑫王真辉田维敏史敏晶
卢亚莉 张世鑫 王真辉 田维敏 史敏晶

摘 要:通過割胶,巴西橡胶树树皮中收集的胶乳被制成天然橡胶。橡胶树树皮的结构在天然橡胶的生产中具有重要地位。死皮是影响天然橡胶产量的重要因子之一,目前对死皮橡胶树树皮结构系统的研究较少。以无性系‘RY7-33-97健康树、三级死皮树和五级死皮树树皮为研究材料,利用光学显微技术,对不同级别、不同死皮程度树皮中的乳管、筛管、石细胞、单宁等的形态结构等进行了研究。结果表明:随着死皮程度的增加,乳管排列逐渐紊乱,不易统计列数;膨大乳管的列数显著增多,从最早出现的砂皮层逐步深入至水囊皮;乳管染色变浅为黄色,甚至出现中空;与直径高达40 ?m的膨大乳管相比,有效乳管的直径通常不足20 ?m,且死皮严重情况下,其直径会进一步减小。筛管的直径在死皮时会发生减小,尤其是树皮不排胶的部位尤为明显。随死皮程度增加,石细胞不仅数量增多,并且形成的位置明显内移,甚至可深达水囊皮,靠近形成层。单宁细胞在树皮中数量随死皮程度增强而逐渐增多,且在水囊皮等幼嫩组织中增加较明显。本研究对比健康树,分析了三级和五级死皮树树皮中多种组织的结构特征,旨在为阐明橡胶树死皮的发生机理提供依据,为死皮防治技术的研发奠定基础。
关键词:橡胶树;死皮;树皮结构;乳管;单宁
中图分类号:S794.1 文献标识码:A
Bark Structure of the Clone ‘RY7-33-97 with Different Level of Tapping Panel Dryness (TPD) in Rubber Tree (Hevea brasiliensis Muell. Arg.)
LU Yali, ZHANG Shixin, WANG Zhenhui, TIAN Weimin, SHI Minjing*
Rubber Research Institute, Chinese Academy of Tropical Agricultural Sciences / Key Laboratory of Biology and Genetic Resources of Rubber Tree of Ministry of Agriculture and Rural Affairs / State Key Laboratory Incubation Base for Cultivation and Physiology of Tropical Crops Rubber Research Institute, Haikou, Hainan 571101, China
Abstract: Natural rubber is extracted from latex that collected from the bark of rubber trees by tapping. The structure of bark plays an important role in the production of natural rubber. Tapping panel dryness (TPD) is one of the key factors to affect yield of rubber trees, but the mechanism of TPD was still less known and the change of bark structure of TPD was rarely studied. By using the optical microscopy technology, the morphological structure and location of laticifer, sieve tube, tannin cell and stone cell in the bark of ‘RY7-33-97 with different level of TPD were studied. Compared to the healthy rubber tree, the bark structure in the third and the fifth grade of TPD tree changed obviously. Accompanying the increase of TPD extent, the number of laticifer lines was difficult to be counted for the disordered arrangement. The swell laticifer appeared gradually inward to cambium from sandy skin. Besides the abnormal swelling and light dying, hollow laticifer also was observed. The diameter of swell laticifer was up to 40 ?m, but the effective laticifer usually was short of 20 ?m and even smaller when TPD aggravated. The diameter of sieve tube decreased obviously under the condition of non-latex flow. Stone cells formed largely in the yellow skin and even went deeply into the water sac skin and the cambium in TPD rubber tree. The number of tannin cells increased obviously when TPD happened, especially in the tender tissue such as water sac skin. This study would lay a foundation for illu-minating the mechanism of TPD and provide a theoretical basis for developing the prevention technology of TPD in rubber trees.
Keywords: Hevea brasiliensis Muell. Arg.; tapping panel dryness (TPD); bark structure; laticifer; tannin
DOI: 10.3969/j.issn.1000-2561.2021.07.014
巴西橡胶树(简称橡胶树)是目前唯一生产具有商业价值天然橡胶的栽培作物[1-2]。天然橡胶作为重要的工业原材料和战略资源,其来源——橡胶树的种植在世界经济发展中占有很重要的地位。我国橡胶树种植面积受限,提高单位面积产量是当前天然橡胶研究中的首要任务。我国胶园中橡胶树死皮现象频发,死皮发生率高达25%~ 50%[3],据估计,死皮导致的干胶损失占年产的12%~14%[4],严重降低了天然橡胶的单产,同时造成了高达上亿元的经济损失[5]。橡胶树死皮即割面干涸病(tapping panel dryness, TPD),即乳管丧失部分或全部产胶能力,表现为割线排胶减少甚至完全停止排胶[6]。为阐明死皮发生发展的机理,人们对其生理学、病理学、组织学以及分子机制等方面展开了广泛的研究[4-11],并研发了死皮防治技术[12-13]。关于橡胶树死皮组织学方面,郝秉中等[9]、吴继林等[10]认为乙烯利过度刺激促进了树皮快速衰老,诱发了死皮的发生;何哲等[11, 14]对耐刺激、晚熟品種‘PR107死皮不同症状割线下方5 cm处的树皮显微结构特征进行了研究。迄今为止,死皮的发生机制仍不清楚。相对健康橡胶树形态结构学的研究,不同品系、不同级别死皮橡胶树树皮结构特征的研究也相对较少。乳管是合成和贮存天然橡胶的组织,最主要的产胶乳管存在于树皮之中。巴西橡胶树树皮从外到内在生产上通常分为粗皮、砂皮外层、砂皮内层、黄皮和水囊皮,产胶乳管则主要分布在砂皮内层和黄皮层中,水囊皮中的乳管主要是未成熟的幼嫩乳管。对树皮乳管在死皮发生中出现的变化进行全面研究是阐明死皮机制研究中的重点。筛管作为运输有机营养物质的主要组织,在橡胶树的正常产排胶中也起到重要作用[15]。单宁细胞和石细胞与橡胶树树皮的抗性防御以及衰老有密切关系[16-17]。因此,本研究主要针对这些组织,以在我国广泛种植的‘RY7-33-97为材料,对健康树、三级死皮树和五级死皮树割线上不同排胶特征部位的形态结构进行了详细研究,旨在为阐明死皮发生发展机制提供依据,为预防和治疗死皮的研发工作奠定基础。
1 材料与方法
1.1 材料
定植于中国热带农业科学院儋州二队试验场中的橡胶树无性系‘RY7-33-97,选取其中长势大致相同的健康树、三级死皮树(即割线死皮长度占总割线长度的1/4~1/2,割线上出现正常排胶部位、点状排胶部位以及完全不排胶的部位)和五级死皮橡胶树(即割线死皮长度占总割线长度的3/4乃至全线死皮,割线上只有点状排胶和完全不排胶2种情况)各3株。采用3天1刀,1/2S的常规割胶制度,分别采集健康树(NL)、三级死皮树正常排胶部位(TF)、点状排胶部位(TP)和不排胶部位(TNF)割线处的树皮以及五级死皮树点状排胶部位的树皮(FP)和不排胶部位(FNF)。停排后用1.4 cm的打孔器采集树皮,深至木质部,沿垂直于树干的方向将材料分割成2块,分别放入80%乙醇和4%多聚甲醛的固定剂中固定,制备石蜡切片。
试剂和仪器:制备石蜡切片所需的二甲苯、乙醇、正丁醇、冰醋酸为广州化学试剂厂国产分析纯,多聚甲醛为Sigma产品,石蜡为生物组织切片专用。切片机为德国Leica RM2235型旋转切片机,成像显微镜为德国Leica DMLB型光学显微镜。
1.2 方法
1.2.1 乳管的观察 参照金华斌等[18]的方法。树皮材料分割后利用80%乙醇固定,并经I-Br处理液特殊变性染色,乙醇梯度脱水,正丁醇透明,石蜡包埋。切片厚度10 ?m,固绿染色,中性树胶封片。
1.2.2 单宁积累的观察 参照史敏晶等[16]的方法,树皮材料分割后利用4%多聚甲醛固定,磷酸缓冲液浸洗后,乙醇梯度脱水,二甲苯透明,石蜡包埋。石蜡厚度10 ?m,二甲苯脱蜡后,汞-溴酚蓝染色,中性树胶封片。参照Shi等[19]的方法,荧光定位分析单宁的自发荧光。
1.3 数据处理
对于乳管观察的切片,每个切片样品中分别选取10个筛管和乳管细胞,沿与韧皮射线平行的方向分别测定其直径大小;乳管列数的计数,根据曾霞等[20]的乳管总列数统计方法。数据采用Excel 2010、Original 8.0软件分析并绘制图形;利用AI和PS软件进行图形的组合。
2 结果与分析
2.1 死皮发生过程中乳管的变化
2.1.1 乳管细胞形态结构的变化 经碘-溴染色后,正常乳管细胞被染色呈棕黑色,出现异常的乳管则染成了黄褐色并开始膨大或者出现中空。健康树中的乳管基本都是正常乳管,染色深,排列紧密而且整齐;少数出现膨大异常,这些异常乳管主要分布在砂皮外层,这可能与该部位的乳管开始出现正常的自然衰老有关(图1A)。
在三级死皮树中,正常排胶部位的树皮结构与健康树基本一致,仅在砂皮外层出现少量膨大的异常乳管(图1B)。点状排胶部位的树皮中膨大乳管的列数明显增加;黄皮层的乳管内出现大量中空,并有少数膨大乳管细胞出现(图1C)。可见,出现点状排胶时,树皮中的乳管已经有了比较明显的异常。膨大乳管通常被染色成黄褐色,基本上丧失了产胶能力,随着死皮程度的增加,这种乳管的数量明显增多。不排胶的树皮中正常乳管进一步减少,主要分布在黄皮层内侧以及水囊皮之中,黄皮层的外侧以及砂皮层中分布的基本都是膨大的、染色浅的异常乳管(图1D)。
在更为严重的五级死皮树中,割线上已经不存在正常排胶的部位。点状排胶部位树皮的黄皮层和砂皮内层基本被膨大的异常乳管占据,能产胶的正常乳管很少;幼嫩的水囊皮中也开始出现膨大乳管(图1E);在不排胶部位的树皮中,除水囊皮外,基本观察不到正常的乳管(图1F),这种结构的异常与不排胶的生理现象相对应。
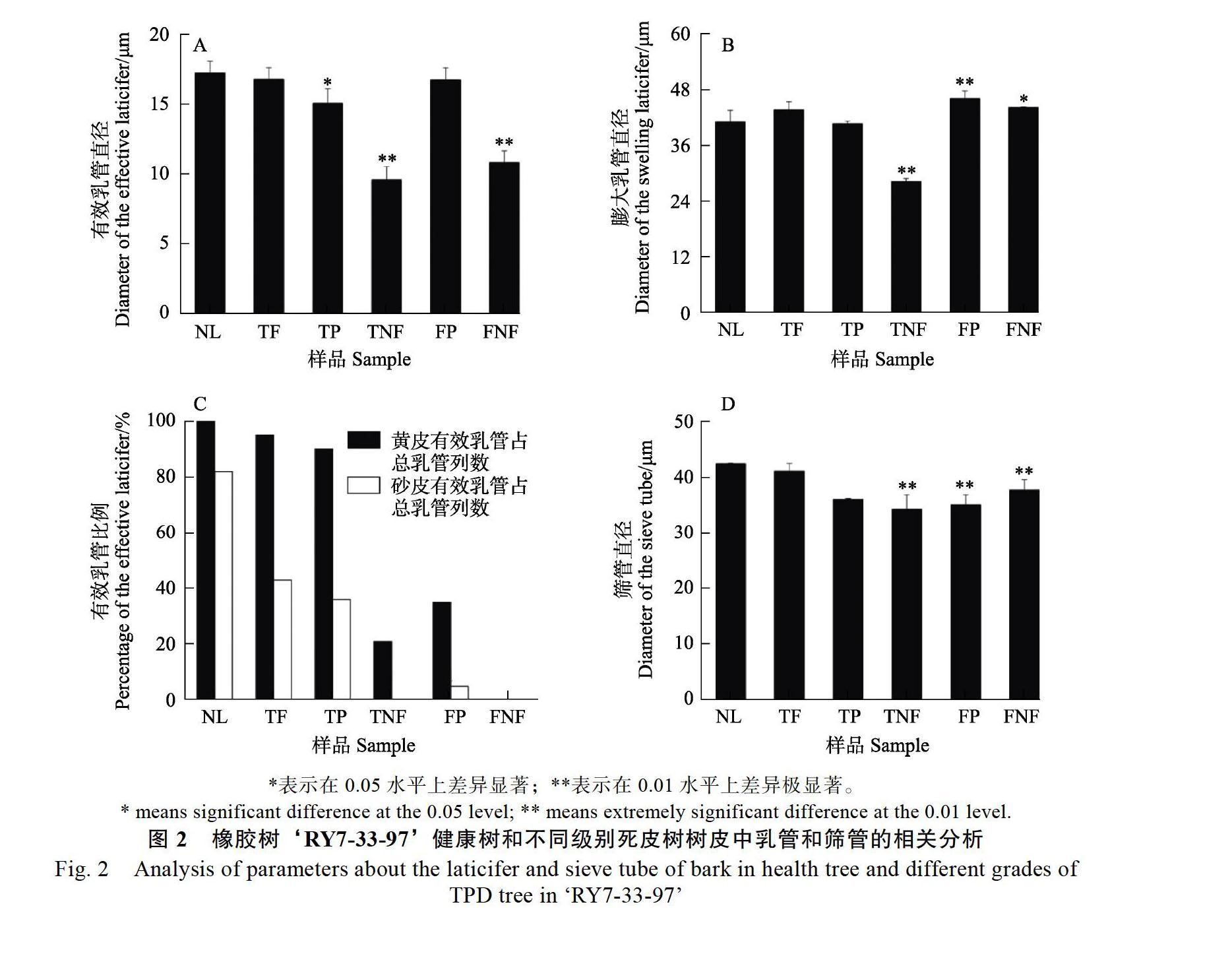
2.1.2 乳管直径的变化 利用统计软件,对不同部位的正常乳管和膨大乳管的直径进行分析。结果表明,正常有效乳管的直径通常低于20 ?m。在健康树和三级死皮树正常排胶部位中,直径分别约为18 ?m和16 ?m;在三级和五级死皮树中的点状排胶情况下,正常乳管直径分别约为
14 ?m和18 ?m;但在完全不排胶的部位,三级死皮树和五级死皮树中统计到的少数正常乳管的直径分别约为9 ?m和11 ?m,极显著小于健康树中的正常乳管的直径(P<0.01)(图2A)。
膨大乳管主要出现在死皮树中,在健康树砂皮外层中偶有出现。经统计,膨大乳管直径可高达40 ?m,甚至更大。健康树和三级死皮正常排胶部位中膨大乳管直径均约为43 ?m;三级和五级死皮树中点状排胶部位观察到的膨大乳管直径分别约为37 ?m和46 ?m,而完全不排胶部位的膨大乳管直径则分别约为27 ?m和44 ?m(图2B)。由此可见,三级死皮树中膨大乳管随着死皮程度的增加其直径有所减小,但五级死皮树中膨大乳管直径变化不明显。
2.1.3 有效乳管比例的变化 对砂皮层和黄皮层2个主要产胶部位中有效乳管所占的比例进行分析(图2C)。结果表明,在健康树砂皮层中有效乳管的比例约82%,出现死皮症状后该比值降低极为明显。外观表现仍正常的三级死皮树正常排胶部位中的有效乳管比例仅为43%,明显低于健康树;三级死皮树点状排胶部位砂皮层中有效乳管占比为36%,但不排胶部位砂皮层中则基本观察不到有效乳管列,占比计为0%。五级死皮树点状排胶部位砂皮层中的有效乳管比例仅5%,远低于三级点状排胶中36%的占比;不排胶部位砂皮层中有效乳管比例为0%。
黄皮层位于砂皮层的内侧,出现异常乳管的时期晚于砂皮层。在健康树中,黄皮层中有效乳
管比例为100%,即尚未出现膨大乳管;三级死皮树中,正常排胶部位和点状排胶部位的有效乳管比例均高达90%以上,可见此时黄皮层受死皮的影响还较小,但在不排胶部位有效乳管比例急剧降至21%,受死皮的影响已经很明显;在五级死皮树中,黄皮层中有效乳管的比例下降明显,点状排胶部位的比例仅35%,不排胶部位则为0%。
由以上结果可见,即使是外在排胶表现相似,但在不同级别的死皮树中,对应的乳管形态结构、直径大小以及其他特征都存在差异,五级死皮树乳管的异常明显比三级死皮树严重。
2.2 筛管、韧皮射线以及石细胞的变化
筛管是纵向输导有机营养物质的主要组织,主要分布在水囊皮。‘RY7-33-97健康树中筛管的直径较大,高达41 ?m;三级死皮树中,从正常排胶、点状排胶到最终完全不排胶,随着死皮症状的加重,筛管直径在逐渐减小,分别为38、36、33 ?m;五级死皮树的点状排胶和不排胶部位对应的筛管直径分别为35 ?m和37 ?m。总体来看,死皮树中筛管直径减小(图2D)。另外,筛管分布的范围在减少,即水囊皮的厚度在变小,这些变化导致韧皮部的纵向输导能力出现明显的减弱。
韧皮射线是树皮中横向运输营养物质的主要组织,通常有规律地从水囊皮延伸至砂皮内层,但到达砂皮外层后往往由于大量石细胞团的出现,导致射线排列扭曲以及中断,最终消失(图1)。比较健康树和三级、五级死皮树不同部位可知,随着死皮程度增加,射线排列开始紊亂,尤其是五级死皮树不排胶的部位,很难观察到清晰的射线。
石细胞作为橡胶树树皮中重要的厚壁组织,主要分布在砂皮层,起支持和保护作用(图1A)。在死皮发生后,石细胞的数量增多,并且逐渐内移至黄皮层甚至水囊皮(图1B~图1F)。在五级死皮树中,尤其是完全不排胶部位,石细胞分布更为深入而广泛,甚至已深至形成层附近(图1E,图1F)。由此认为,石细胞过早出现在水囊皮等幼嫩组织中也是树皮死皮的特征之一。
2.3 单宁的变化
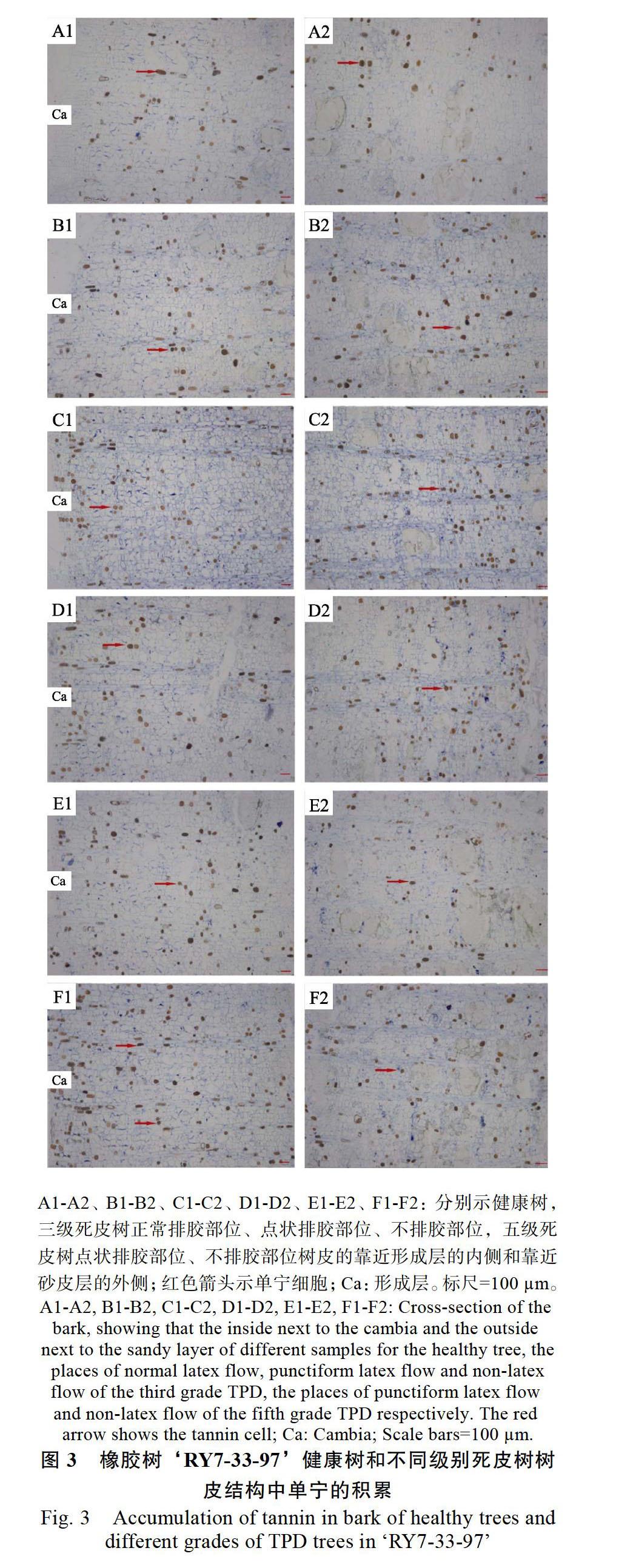
橡胶树树皮中含有大量的单宁细胞。多聚甲醛可以固定组织中的单宁,不需要染色便能在光镜下观察到棕红褐色的单宁物质[9];在荧光显微镜下,单宁自发红色荧光,利用这些单宁的特性,分析‘RY7-33-97的健康树、三级和五级死皮树不同程度死皮部位的树皮内单宁的积累情况。光学显微镜下,汞-溴酚蓝染色后蛋白质类物质被染色成鲜蓝色[13, 19],与呈红褐色的单宁物质形成明显的对比。在健康树中,单宁细胞较少,零星分散在从水囊皮到砂皮外层区域的射线薄壁细胞和其他韧皮薄壁细胞中(图3A1,图3A2)。三级死皮树中正常排胶部位树皮和健康树树皮中单宁积累的程度相差不大,单宁细胞分布仍然较为稀疏(图3B1,图3B2),但随着死皮程度的增加,单宁细胞密度增大,且逐步深入分散至水囊皮甚至靠近形成层的幼嫩组织中(图3C1,图3C2,图3D1,图3D2)。在五级死皮树中,单宁细胞分布最密集的区域主要是水囊皮(图3E1,图3E2,图3F1,图3F2)。可见,随着死皮程度的增加,单宁细胞不断增加,表明死皮可诱导单宁的产生,这可能与单宁的抗氧化、抗衰老等功能有关。
荧光显微镜下单宁物质经绿光激发后自发红色荧光。由结果可知(图4),健康树以及三级死皮树的正常排胶部位中,红色荧光少而弱,并且主要靠近树皮的外侧(图4A1,图4B1);随着死皮程度的逐渐增加,红色荧光明显增强,表明单宁物质的积累明显增多,并且分布位置也开始明显靠近树皮内侧(图4C1,图4D1,图4E1,图4F1)。由合并图中的明场可以清晰地看到单宁积累的位置,既可以在普通薄壁细胞中,也可以在射线细胞中;健康树皮中单宁很少出现在水囊皮以及形成层附近(图4A2),而死皮树尤其在发生严重的情况下,即使是形成层区、水囊皮等幼嫩组织也开始大量积累单宁(图4B2,图4C2,图4E2,图4F2)。自发荧光定位与化学染色显示的结果基本一致,展现的规律相同,但不同死皮程度橡胶树中荧光显示的结果对比更为鲜明。
3 讨论
‘RY7-33-97作为早熟高产品种,其干胶含量平稳、产量高、抗风力强,在我国植胶园中被广泛推广种植。作为主要栽培新品种,其在我国累积推广面积高达16.67万hm2[21]。但‘RY7-33-97不耐受乙烯利刺激,容易被乙烯利诱发死皮。研究‘RY7-33-97的死皮发生机理不仅为全面解析橡胶树死皮机制提供基础,对于天然橡胶生产中维持高产、稳产也具有重要意义。
发生死皮的橡胶树是随机的,在取材上往往难以保证研究材料的一致。本研究在选取试验材料时确保了每个样品至少3株重复,并且重复的3株树的排胶量及排胶时间相近,同时健康树、三级死皮树、五级死皮树3组一共9株树的树围基本一致,以此保证试验的可重复性和准确性,排除了因生长发育差异带来的乳管列数差别大而不利于后续分析的问题。
通过不同级别死皮树膨大乳管出现的位置以及比例,可以得出死皮的发生尤其是乳管的改变是从外发展到内部的,首先是砂皮层乳管异常,然后才发展到黄皮层,甚至水囊皮,由于膨大乳管中的胶乳发生了原位凝固而不能排胶,所以最早的表现就是排胶线出现明显的内缩,然后才是排胶是否连续。乳管的原位凝固和封堵可能对橡胶树本身而言是一种有效的逆境防护。三级死皮树在正常排胶和点状排胶部位黄皮层的正常乳管仍然占有极高的比例,在90%以上,但随后随着死皮程度的增加该比值迅速下降。因此,研究认为三级死皮树中点状排胶现象的出现可能是一个分界线,随之可能出现死皮的明显加重,但这一观点还需要对更多不同级别的死皮树进行分析后才能确定。
筛管和射线作为韧皮部的主要纵向和横向运输组织,在死皮发生后,筛管的直径减少并且其所在的水囊皮在韧皮部的比例降低;而射线出现紊乱,这些变化显然会导致橡胶树树皮运输营养物质的能力减弱,而橡胶树乳管合成胶乳需要消耗大量营养,运输不力显然是不利于乳管的正常代谢以及胶乳的合成,最终可能会进一步加剧死皮。
石细胞和单宁细胞作为支持和防御的组织结构,通常先出现在橡胶树树皮的外围,也是一种组织出现衰老的表现。死皮发生的过程中,‘RY7-33-97和其他品系一样,同样出现石细胞和单宁明显增多的现象,主要表现出从砂皮层向水囊皮内移的发展趋势[9, 11]。从统计数据和单宁荧光显示结果可知,三级死皮正常排胶部位的单宁细胞还较少,在水囊皮的分布也极少,但点状排胶出现后,单宁细胞在水囊皮的分布即有明显的增加。水囊皮出现大量衰老相关的组织,意味着死皮的发生已经较为严重。在实际生产中胶工通常将树皮颜色加深甚至显示出棕紅色作为判断死皮发生的依据之一,即与大量单宁的形成有关。史敏晶等[16]研究表明,橡胶树树皮组织在受到乙烯利刺激、机械伤害、JA刺激等处理时都能形成大量单宁,表明单宁受多种因素尤其是逆境的诱导。植物在逆境形成大量的单宁应与单宁的防御功能有关。单宁是一种对蛋白质具有凝絮功能的次生代谢物[22],而胶乳中含有大量的蛋白质,那么单宁是否具有促进胶乳凝固以及乳管原位凝固的作用?单宁物质在死皮发生中具体起到何种作用目前尚不清楚,具有进一步研究的价值。
参考文献
[1] Priyadarshan P M, Goncalves P de S. Hevea gene pool for breeding[J]. Genetic Resources and Crop Evolution, 2003, 50(1): 101-114.
[2] 袁红梅, 洪 灏, 黄 惜. 巴西橡胶树产排胶机理的研究进展[J]. 分子植物育种, 2015, 13(5): 1151-1156.
[3] 邹 智, 杨礼富, 王真辉, 等. 橡胶树“死皮”及其防控策略探讨[J]. 生物技术通报, 2012(9): 8-15.
[4] Chen S C, Peng S Q, Huang G X, et al. Association of decreased expression of a Myb transcription factor with the TPD (tapping panel dryness) syndrome in Hevea brasiliensis[J]. Plant Molecular Biology, 2003, 51(1): 51-58.
[5] 覃宝祥, 胡新文, 邓晓东, 等. 橡胶树死皮病的形成及其机制[J]. 植物生理学通讯, 2005, 41(6): 827-830.
[6] 袁 坤, 徐智娟, 王真辉, 等. 橡胶树胶乳死皮相关蛋白的鉴定及分析[J]. 西北林学院学报, 2012, 27(6): 105-109, 127.
[7] Putranto R A, Herlinawati E, Rio M, et al. Involvement of ethylene in the latex metabolism and tapping panel dryness of Hevea brasiliensis[J]. International Journal of Molecular Sciences, 2015, 16(8): 17885-17908.
[8] 袁 坤, 周雪梅, 李建辉, 等. 死皮防治剂对死皮橡胶树胶乳生理的影响[J]. 湖北农业科学, 2011, 50(17): 3570-3572.
[9] 郝秉中, 吴继林, 云翠英. 乙烯利刺激割胶对巴西橡胶树树皮结构的影响[J]. 热带作物学报, 1980, 1(1): 61-65, 110.
[10] 吴继林, 谭海燕, 郝秉中. 乙烯利过度刺激采胶诱导巴西橡胶树割面干涸病的研究[J]. 热带作物学报, 2008, 29(1): 1-9.
[11] 何 哲, 王真辉, 林位夫, 等 .巴西橡胶树不同死皮症状的解剖结构特征[J]. 吉林农业, 2010, 243(5): 40-41.
[12] 周 敏, 王真辉, 李 芹, 等. 橡胶树死皮防控试验[J]. 热带农业科学, 2016, 36(12): 52-55.
[13] 袁 坤, 白先权, 冯成天, 等. 死皮康复营养剂恢复橡胶树热研7-33-97死皮植株产胶能力的效果分析[J]. 热带作物学报, 2017, 38(7): 1253-1259.
[14] 何 哲. 橡胶树死皮发生过程不同排胶现象的树皮解剖学特征[D]. 海口: 海南大学, 2010.
[15] 郝秉中, 吴继林, 云翠英. 巴西橡胶树有输导功能的韧皮部与采胶的关系[J]. 植物学报, 1980, 22(3): 227-231, 310.
[16] 史敏晶, 吴继林, 郝秉中, 等. 巴西橡胶树树皮单宁细胞结构和发育[J]. 林業科学, 2014, 50(8): 45-52.
[17] 史敏晶, 李 言, 王冬冬, 等. 巴西橡胶树树皮厚壁组织结构和发育研究[J]. 热带作物学报, 2016, 37(2): 311-316.
[18] 金华斌, 田维敏, 胡彦师, 等.橡胶树魏克汉种质苗期初生乳管分化能力研究[J]. 热带作物学报, 2017, 38(10): 1769-1776.
[19] Shi M, Li Y, Deng S, et al. The formation and accumulation of protein-networks by physical interactions in the rapid occlusion of laticifer cells in rubber tree undergoing successive mechanical wounding[J]. BMC Plant Biology, 2019, 19(4), https://doi.org/ 10.1186/s12870-018-1617-6.
[20] 曾 霞, 李维国, 胡彦师, 等. 5龄热试99橡胶树乳管特征及其与生长和产量的关系[J]. 热带作物学报, 2006, 27(3): 1-4.
[21] 仇 键, 校现周, 高宏华, 等. 中龄PR107和热研7-33-97橡胶树“6天1刀”割制的刺激技术及适应性研究初报[J]. 热带作物学报, 2020, 41(3): 474-481.
[22] 王 洁. 多酚-蛋白质相互作用的影响因素及其功能特性研究进展[J]. 河南工业大学学报(自然科学版), 2012, 33(3): 91-96.
责任编辑:谢龙莲
