延龄草总皂苷体内外抗结肠癌活性的作用
2021-08-23陈静文陈先勇赵龙飞柳蔚李世刚喻玲玲
陈静文 陈先勇 赵龙飞 柳蔚李世刚喻玲玲*
[1.三峡大学医学院,湖北 宜昌443002; 2.三峡大学人民医院(宜昌市第一人民医院) 病理科,湖北宜昌443000; 3.天然产物研究与利用湖北省重点实验室,湖北 宜昌443002; 4.国家中医药管理局中药药理(肿瘤) 科研三级实验室,湖北 宜昌443002]
结肠癌是消化系统中最为常见的恶性肿瘤之一[1],严重威胁人类健康。化疗仍是治疗结肠癌的主要手段,但化疗药物严重的不良反应等问题使其临床应用受到很大的限制。因此寻找能高效杀伤肿瘤细胞同时毒副作用小的药物,是当今抗肿瘤新药研发的热点。
延龄草Trillium tschonoskiiMaxim.是土家族四大名药之一,别名“头顶一颗珠、芋儿七”。其味甘性温,有消痛去肿、止血解毒、安神活血等功效,主治肺痈疮肿、疔疮肿毒、头痛眩晕、神经衰弱等疾病,因其果实呈圆球形生长在上部故得名“天珠”。而其根茎下方生多数细根,加工成药材时常将其编扎在根茎之外方,形成球状,又因其生长在地下,又称“地珠”[2]。延龄草总皂苷是其主要的化学成分[3]。研究表明,延龄草总皂苷(Trillium Total Saponins,TTB)对结肠癌、肺癌、宫颈癌等均有显著抑制作用[4]。RAS是目前所知最保守的癌基因之一,参与细胞生长、增殖、发育及分化调控等重要生理过程,也在细胞恶性转化中起重要作用。RAS 蛋白的持续活化可引起细胞快速增殖和抗凋亡信号亢进,由此导致肿瘤的发生和发展。Hans Raskov 等[5⁃6]研究发现,结肠癌的发生与K⁃ras、APC和P53 基因突变高度相关。目前,已有许多研究证实延龄草总皂苷可以显著抑制结肠癌细胞增殖[7],诱导结肠癌细胞凋亡[8],抑制结肠癌细胞转移[9],但其可能的分子机制尚未阐明。因此,本课题组观察了延龄草总皂苷对裸鼠皮下肿瘤的抑制作用以及其对结肠癌CT26细胞增殖的抑制作用,探讨其可能发生的作用机制,以期为结肠癌的临床治疗提供基础理论支撑。
1 材料
1.1 药材 本实验所用中药延龄草地珠采摘于神农架林区,经三峡大学生物技术研究中心陈发菊教授鉴定为延龄草地珠Trillium tschonoskiiMaxim.,植物标本保存于三峡大学天然产物研究与利用湖北省重点实验室(编号TT200509SNJ)。
1.2 试剂 甲醇购自天津市天力化学试剂有限公司(批号20180708);乙醇购自上海沪试化学试剂有限公司(批号20190123);石油醚购自天津市科密欧化学试剂有限公司(批号20161207);乙酸乙酯购自广东西陇科学股份有限公司(批号160805);正丁醇购自广东华大化学试剂有限公司(批号20130810);RPMI⁃1640 培养基购自武汉普诺赛生命科技有限公司(批号WHO1111803SP);胰蛋白酶购自杭州吉诺生物有限公司(批号20180806);胎牛血清(批号1856032)购自美国gibco 公 司;二甲基亚 砜(Dimethyl sulfoxide,DMSO)购自国药集团化学试剂有限公司(批号20160704);MTT 试剂购自武汉塞维尔生物科技有限公司(批号180909);BCA 试剂盒(批号RC230516)购自美国Thermo Scientific 公 司;β⁃actin 抗 体(批号181109)购自武汉塞维尔生物科技有限公司;RAS 抗体(批号5)、Akt 抗体(批号20)、p⁃Akt 抗体(批号23)、ERK1/2 抗体(批号21)、p⁃ERK1/2 抗体(批号17)均购自美国CST 公司。
1.3 仪器 EYELAN⁃1100型旋转蒸发仪购自上海爱郎仪器有限公司;电子天平购自上海天平仪器厂;全波长酶标仪购自美国Thermo Electron 公司;MCD175型CO2培养箱购自日本Sanyo 公司;Eppendorf centrifuge 5804R型低温高速离心机购自德国Eppendorf 公司;超净台购自苏州安泰空气技术有限公司;微型瞬时离心机、免疫印迹仪,凝胶成像仪购自美国Bio⁃Rad 公司。
1.4 瘤株 小鼠CT26细胞购自上海柯雷生物技术有限公司。
1.5 动物 BALB/c⁃nu 雌性鼠与雄性鼠各20 只,SPF 级,体质量18~20 g,由湖北省实验动物研究中心供应,由三峡大学动物实验中心饲养,实验动物使用许可证编号为SYXK(鄂)2011⁃0061。
2 方法
2.1 延龄草总皂苷制备 取延龄草地珠于48 ℃恒温干燥,粉碎后过60 目筛,得到6.4 kg 原药材粉末。以甲醇为溶剂,利用多功能强制渗漉罐(常压,65 ℃)提取,每2 h提取1 次,反复3 次。提取液过滤后用旋转蒸发仪减压浓缩(温度48 ℃),干燥得深棕色醇提物浸膏共2.4 kg。甲醇提取浸膏和蒸馏水以1∶1 比例充分混匀,依次用石油醚、乙酸乙酯和正丁醇萃取,萃取液减压浓缩(0.1 mPa,50 ℃),真空冷冻干燥得各个萃取物。其中正丁醇萃取部分浸膏723 g,为延龄草总皂苷。经比色法测定延龄草中总皂苷的含有量为19.78%。
2.2 建立移植瘤小鼠模型 BALB/c⁃nu 鼠在三峡大学动物实验中心适应性饲养一周后开始实验(体质量18~20 g)。取对数生长期结肠癌CT26细胞,台盼蓝拒染显示活细胞>95%,调整细胞浓度至4×106/ L,制成细胞悬液,于右腋皮下接种0.2 mL 形成实体瘤。待肿瘤大小约为100 mm3后将荷瘤小鼠随机分为对照组、5⁃FU组及高、中、低浓度组,每组8 只。模型组给予生理盐水灌胃(0.2 mL),低、中、高剂量组分别用延龄草总皂苷水溶液(5、10、15 mg/kg)灌胃。每天给药1 次,共9 d。
2.3 肿瘤抑制率的计算 每2 天测量1 次体质量,观察给药组和对照组荷瘤小鼠体征和体质量变化情况,并绘制出体质量变化图。给药第9 天,处死荷瘤小鼠,剥取瘤组织并称取质量,计算肿瘤抑制率。抑瘤率=[(对照组平均瘤重-给药组平均瘤重)/对照组平均瘤重]×100%。
2.4 病理切片HE 染色 荷瘤小鼠称定质量处死后,剥离瘤体组织,剪取1 cm×1 cm组织固定于10% 甲醛溶液中,常规石蜡包埋切片,苏木精⁃伊红染色法(hematoxylin⁃eosin staining,HE)染色,镜下观察肿瘤组织病理学变化。
2.5 RT⁃PCR 检测瘤体组织细胞RAS基因表达 称取150 mg瘤组织置于干燥的玻璃匀浆器内,加1 mL TRIzol,充分研磨,提取RNA,利用反转录试剂盒将其反转为cDNA,然后RT⁃PCR 的方法检测RAS基因的表达。采用2-△△CT法分析实验组和对照组细胞之间RASmRNA 表达的相对差异。引物序列见表1。

表1 引物序列
2.6 MTT 法检测延龄草总皂苷对CT26 体外增殖抑制作用 将处于对数生长期的结肠癌CT26细胞以5×104/mL 接种于96 孔板内,每孔100 μL,置于5% CO2、37 ℃恒温培养箱中培养24 h。分别加入含有不同浓度的延龄草总皂苷(0、3.125、6.25、12.5、25、50 μg/mL)的培养液100 μL,溶剂对照组仅加入含有0.054% DMSO 的培养液,每孔设置3个平行复孔,细胞继续培养24 h。然后取出培养板,每孔加入5 g/L 的MTT 溶液20 μL,继续孵育4 h 后,弃去留存的孵育液,每孔加入150 μL DMSO 置摇床低俗振荡10 min使得结晶充分溶解,酶标仪上检测各孔490 nm 处吸光度值(OD)。计算细胞增殖率=[(OD给药组-OD对照组)/OD对照组]×100%。
2.7 Western blot 测定结肠癌细胞中RAS、Akt、p⁃Akt、ERK1/2、p⁃ERK1/2 的表达以及瘤体组织细胞RAS 蛋白的表达 收集对照组和延龄草总皂苷处理各组结肠癌CT26细胞,用细胞刮收集后加入RIPA细胞裂解液提取总蛋白,用BCA 试剂盒进行蛋白定量。蛋白变性后进行聚丙烯酰胺凝胶电泳,转膜。聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)膜用5%脱脂牛奶室温封闭1 h,分别加兔抗RAS(1∶ 1 000)、Akt(1∶ 1 000)、p⁃Akt(1∶ 1 000)、ERK1/2(1∶ 1 000)、p⁃ERK1/2(1∶ 1 000)及兔抗β⁃actin(1∶ 1 000)一抗4 ℃孵育过夜,TBST 洗膜后加对应HRP 标记二抗,室温孵育2 h,TBST 洗膜后,进行化学发光。瘤体组织细胞的处理与之类似,取0.1 g 瘤组织放入玻璃匀浆器,加入1 mL RIPA 裂解液,冰上充分研磨,提取蛋白,用相同的方法检测其中RAS 蛋白的表达。
2.8 统计学分析 采用SPSS 22.0 统计软件进行数据统计,Western blot 采用Image J 进行灰度分析。定量数据以()表示,n为样本数量,两组间计量资料的比较采用独立样本t检验。P<0.05 为差异具有统计学意义。
3 结果
3.1 肿瘤抑制率 如表2 所示,5⁃FU组肿瘤抑制率最高为65.15%,延龄草总皂苷高、中、低浓度组也都表现出较好的抑制率,分别为51.22%、34.88%和25.02%,呈一定的浓度依赖趋势。此结果说明延龄草总皂苷对小鼠体内肿瘤的生长有抑制效果。
表2 延龄草总皂苷对小鼠瘤重的影响(, n=8)

表2 延龄草总皂苷对小鼠瘤重的影响(, n=8)
注:与对照组比较,**P<0.01。
3.2 延龄草总皂苷对RAS基因表达的影响 如图2 所示,与对照组相比,延龄草总皂苷的低、中、高浓度组均能降低RAS基因的表达(P<0.05,P<0.01),呈浓度依赖性。因此推测延龄草总皂苷抑制肿瘤的生长与抑制RAS基因表达或者其下游通道基因相关联。
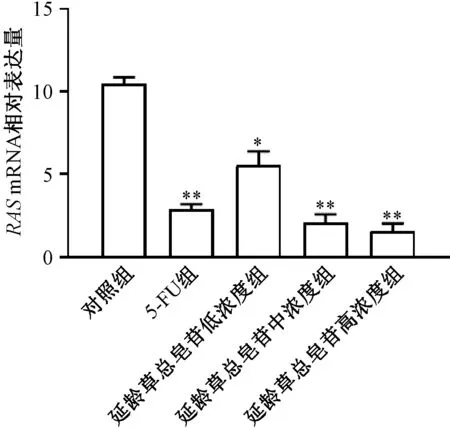
图2 延龄草总皂苷对RAS 基因表达的影响(n=8)
3.3 延龄草总苷作用肿瘤组织及脾组织HE 染色结果 与对照组相比,延龄草总皂苷的低、中、高浓度组均可见肿瘤细胞坏死,尤其以中高浓度组明显,其中可见大片肿瘤细胞坏死。由此说明延龄草总皂苷可促进结肠癌细胞的凋亡(图3)。结果显示多数细胞核显深色,且胞质致密、嗜酸性染色(伊红染色)增强,呈凋亡状态。同时,为了观察延龄草总皂苷对小鼠是否有器质性的影响,对荷瘤鼠的脾脏进行切片。如图4 所示,与对照组相比,各组脾脏切片均未见明显病变,无明显差异,说明延龄草总皂苷对正常组织的毒副作用小。

图3 延龄草总苷作用后肿瘤组织HE 染色(×40)

图4 延龄草总苷作用后脾脏组织HE 染色(×40)
3.4 延龄草总皂苷降低体内RAS 蛋白的表达 如图5 所示,与对照组相比,延龄草总皂苷的低、中、高浓度组均能降低RAS 蛋白的表达(P<0.01),而且呈剂量依赖性。

图5 延龄草总苷对肿瘤组织RAS 蛋白表达的影响(n=8)
3.5 延龄草总皂苷抑制结肠癌CT26细胞增殖 延龄草总皂苷处理体外培养CT26细胞24 h 后,MTT 法分析药物对细胞增殖能力的影响,图6 显示延龄草总皂苷对CT26细胞增殖具有抑制作用,抑制效果随总皂苷浓度的增加而增强。在处理24 h 条件下,延龄草总皂苷作用于CT26细胞的IC50为(10.23±1.02)μg/mL。依据上述结果,后续实验使用终质量浓度分别为5、10 μg/mL 延龄草总皂苷处理的细胞作为实验组,以0 μg/mL 处理的细胞作为对照组。
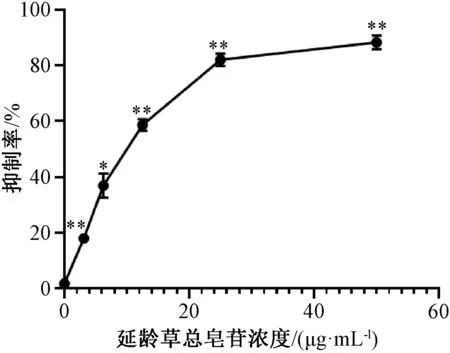
图6 延龄草总皂苷对结肠癌CT26细胞的增殖抑制作用(n=3)

图6 延龄草总皂苷对结肠癌CT26细胞蛋白表达的影响(n=3)
3.6 延龄草总皂苷降低结肠癌CT26细胞RAS 相关蛋白的表达 进一步分析延龄草总皂苷对于结肠癌CT26细胞中RAS/AKT/ERK 信号通路的影响。结果发现延龄草总皂苷的高浓度组(10 μg/mL)、低浓度组(5 μg/mL)均能呈浓度依赖性降低RAS 蛋白以及AKT、ERK1/2 及其磷酸化蛋白的表达。由此推测延龄草总皂苷抑制肿瘤生长与抑制RAS/AKT/ERK 信号通路有关。
4 讨论
RAS 基因在许多肿瘤的发生发展过程中产生重要的作用,参与细胞生长、增殖、发育、分化以及恶性调控等重要的生理过程,同时RAS 蛋白也是表皮生长因子受体信号通路中的重要成员。表皮生长因子受体(epithelial growth factor receptor,EGFR)参与的细胞信号传导通路在结直肠癌原发灶形成中扮演着重要角色,其主要通过丝裂原活化蛋白激酶(RAS⁃RAF⁃ERK)和磷脂酞肌醇激酶(PI3K⁃PTEN⁃AKT)两条途径,将细胞外信号转导至细胞内细胞核,从而引起核内基因转录水平的增加,进而调控细胞的增殖、分化、转化和凋亡等重要的生理过程[10]。目前也有许多针对RAS/RAF/MEK/ERK 这一信号通路的靶向药物出现[11],Jeong 等[12]提出了通过降低蛋白质活性从而干预RAS⁃ERK 信号通路抗结肠癌治疗的一系列新策略。因此阐明延龄草总皂苷是否通过干扰Ras 信号通路发挥其抗结肠癌效果成为本研究的重要内容。
在本研究中,体外细胞实验表明延龄草总皂苷对于体外培养的结肠癌CT26细胞的增殖具有抑制作用,其24 h时IC50值为(10.23±1.02)μg/mL,表明其可能是抗结肠癌的有效成分。体内实验表明延龄草总皂苷有效抑制了裸鼠皮下肿瘤的生长,延龄草总皂苷高、中、低三个浓度也都表现出较好的抑制率,分别为51.22%、34.88% 和25.02%,呈一定的浓度依赖趋势(见表1)。此结果说明延龄草总皂苷体内对肿瘤的生长有抑制效果,与体外细胞实验符合。
结肠癌的发生与RAS 蛋白的持续活化有关,体内实验瘤组织蛋白水平的结果进一步验证其剂量依赖性作用(见结果2),而延龄草总皂苷的高、中、低3个剂量组在mRNA 水平上均能显著降低RAS基因的表达,而且呈剂量依赖性。因此推测延龄草总皂苷抑制肿瘤的生长与抑制RAS基因表达或者其下游通道有关联。随后在体外实验中使用Western blot 法检测细胞中RAS、AKT、ERK1/2 蛋白及其磷酸化蛋白的表达水平进一步证实其作用机制,发现RAS 蛋白、磷酸化AKT 蛋白、磷酸化ERK1/2 蛋白同时呈现浓度依赖性降低趋势。说明延龄草总皂苷在RAS 信号通路的活化过程中下调RAS 信号通路下游分子的表达。HE染色结果同时也提示体内中高剂量组延龄草总皂苷可以促进结肠癌细胞的凋亡。
综上所述,延龄草总皂苷在体内外均具有抗结肠癌药理活性,其机制可能是通过RAS/AKT/ERK 信号通路干扰其下游蛋白的表达,至于其具体的作用靶点还有待进一步研究。本研究有望为抗结肠癌药物开发提供一个新的思路。
