不同倍性云南山茶种子表型及萌发特性研究
2021-08-19李紫倩王兵益
李紫倩, 左 杰, 王兵益, 屈 燕
(1.西南林业大学园林园艺学院,国家林业和草原局西南风景园林工程技术研究中心,云南省功能性花卉资源及产业化技术工程研究中心, 昆明 650224;2.中国林业科学研究院资源昆虫研究所, 云南 昆明 650224)
云南山茶(CamelliareticulataLind1.f.SimplexSealy)为山茶科(Theaceae)山茶属(Camellia)植物,又名腾冲红花油茶、南山茶、油茶果等,是中国特有种,属国家二级重点保护植物,花色丰富艳丽、油脂含量较高,为观赏兼食用植物,开发前景广阔[1-2]。云南山茶主要分布于云南、四川西南部和贵州西部,生长在海拔1 000~3 200 m之间,野生居群具有三个倍性[3-4],即二倍、四倍和六倍体。
由于生态环境恶化,云南山茶野生居群日渐萎缩。目前主要繁殖方法为嫁接繁殖[5],但嫁接技术尚不成熟且受季节影响。对云南山茶萌发速率[6]、耐旱[7]、耐寒[8]及杂交起源[9]、细胞多样性[10]等研究较多,但有关云南山茶种子萌发,仅有王兵益等[6]对云南山茶种子进行了低温沙藏促进萌发的研究,而对三个倍性云南山茶种子具体表型及萌发特性未有进一步研究分析。萌发特性是引种驯化工作前期重要的基础工作,因此,本研究以不同倍性云南山茶种子为材料,观测了不同倍性表型及低温沙藏对云南山茶播种后其种子发芽势和发芽率等萌发指标的影响,以及不同浓度梯度的GA3对高海拔居群种子各项萌发指标的影响,拟解决各倍性表型及萌发特性问题,为云南山茶种质资源保护和栽培应用提供理论依据。
1 材料与方法
1.1 试验材料
供试的3个云南山茶自然居群分别位于四川省盐边县格萨拉园艺场(材料编号YY,下同),四川省会东县马龙乡(ML),云南富民县东村镇(FM),各倍性的地理位置及主要信息见表1。每个倍性居群选择20株样株,株间距50 m以上;在每个样株树冠中上部南向位置采集5~8个蒴果,于2020年9—10月完成采样。蒴果在通风干燥处晾干后将种子取出,放入4 ℃冷藏待用。
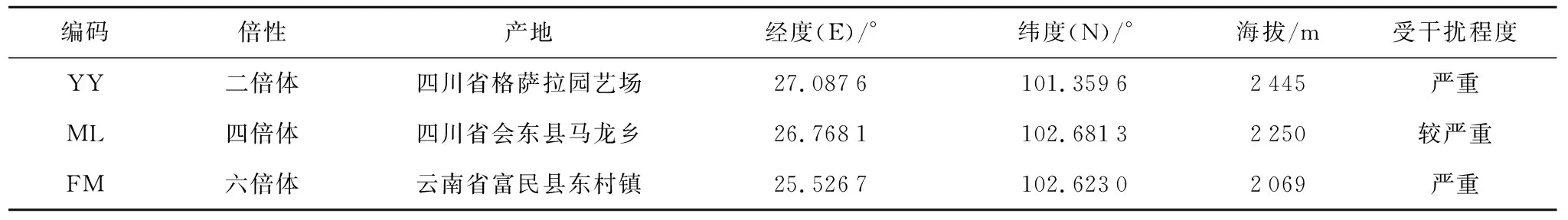
表1 不同倍性云南山茶采集地信息
1.2 试验方法
1.2.1种实表型性状测定
测定指标包括蒴果长度、宽度、厚度和质量以及种子的长度、宽度、厚度和百粒重。从每个倍性随机选择100个蒴果,用游标卡尺分别测定蒴果的最长、最宽及最厚处,测定精度0.1 cm,8次重复并求均值。在每个倍性的每一样株上随机选择100粒种子,用游标卡尺测量其最长、最宽及最厚处,8次重复并计算平均值。
1.2.2种子百粒重测定
参考GB 2772—1999《林木种子检验规程》[12],从测定样品种子中取出100粒,8个重复,各重复分别用千分之一电子秤称重(g),计算均值。
1.2.3种子吸水率测定
参照《1996国际种子检验规程》[13]的方法。将每个倍性测定样品分为3组,每组3个重复,求各组平均值。
1.2.4种子含水量测定
每个倍性随机取出30粒种子,称取质量(m1)后放入80 ℃的烘箱中24 h,取出后冷却至室温再称取各倍性质量(m2),每个倍性3次重复,取均值。
含水量(%)=[(m1-m2)/m1]×100%
1.2.5种子生活力测定
参考《1996国际种子检验规程》[12]方法,采用TTC(2,3,5-氯化三苯基四氮唑)染色后用固定波长分光光度计测定云南山茶种子生活力,每个倍性3次重复,求其平均值。
1.3 种子萌发实验
将3个倍性种子先用0.5%的高锰酸钾消毒1 h,然后用蒸馏水漂洗数遍直至种皮无紫色,之后与细砂及水混匀后分别放在不同塑封袋中,4 ℃沙藏50 d,每7 d检测一次水分状况,干燥时立即洒水即可,达到握土成团,一触即散的状态最佳。对照组在室温条件下贮藏。50 d后将沙藏种子取出与对照组种子放在铺有无菌滤纸的培养盒中,在温度25 ℃,湿度85%,光照强度12.5 μmol·(m2·s)-1的三洋培养箱中进行萌发实验。每个处理重复3次,每次重复均随机选取30粒种子。每天定时观察统计,并将萌发完成的种子移出培养盒。
另对YY(二倍体)云南山茶种子进行0、200、400、600 mg·L-1的GA3浸种处理,0即为清水浸种作为ck。消毒方法同上,浸种时长24 h,萌发条件同上。
1.3.1萌发指标测定
参考李先民等[14]的统计方法,以根长5 mm为萌发标准。分别计算下列萌发指标。
发芽率为种子萌发结束时所有已发芽种子数占供试种子总数的百分比。
发芽率(%)=(已发芽种子数/供试种子总数)×100%。
发芽势为种子发芽达到峰值时统计已发芽种子数与供试种子总数的百分比。
发芽指数=∑Gt/Dt,式中,Gt为在时间t(d)的发芽数;Dt为相应的发芽天数。
平均发芽时间=∑(D×n)/∑n,式中,D为从种子置入培养箱起算的天数;n为相应各天的发芽种子数。
1.3.2数据分析
用Excel软件统计不同倍性的各项测定结果并使用OriginPro 9.0软件进行绘图分析,运用SPSS 25软件进行显著性方差分析。
2 试验结果
2.1 不同倍性云南山茶种子表型差异分析
由表2可知,云南山茶蒴果的长宽比均在0.74~1.02范围内,百粒重和蒴果质量均为YY>ML>FM。Pearson相关性分析表明,海拔与蒴果长宽比呈正相关(r=0.573),与蒴果质量也呈正相关(r=0.647),这与王桔红等[16]的研究结果一致。ANOVA单因素方差分析表明,种子长宽比居群间差异显著(p<0.05),纬度与各倍性种子长度呈负相关(r=-0.992),与蒴果长宽比也为负相关(r=-0.580)。种子含水量为YY>ML>FM,经过LSD多重比较显示,种子含水量间均有显著差异(p<0.05),Pearson相关性分析表明,海拔与云南山茶种子含水量呈显著性正相关(r=-0.763),即随海拔升高种子含水量降低,这与屈燕等[15]的研究结果相同。另外百粒重为YY>ML>FM,组间差异显著(p<0.05);单因素方差分析表明,居群间差异显著(F=6.74,p<0.05)。
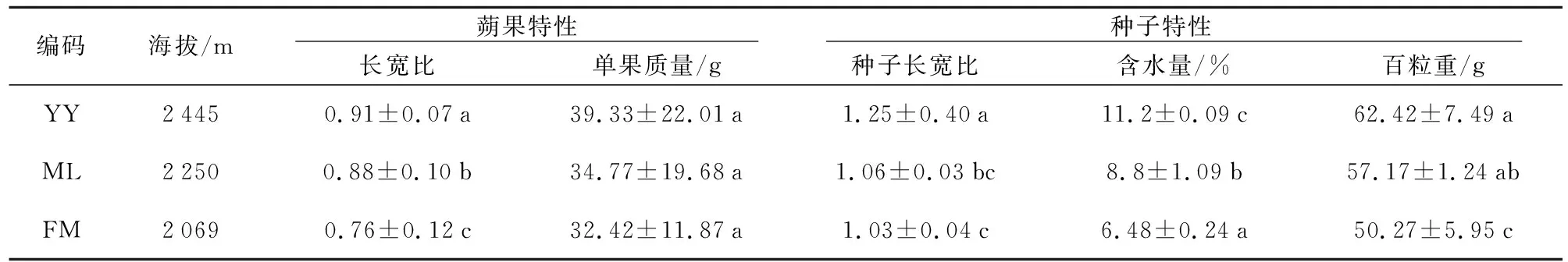
表2 不同倍性云南山茶表型比较
2.2 低温层积处理对不同倍性云南山茶种子萌发的影响
由图1 A可知,对照组整体萌发指标均低于低温处理组,其中3个倍性的平均萌发率为22%,ML的种子发芽势虽不是最高,但发芽率却最高,Pearson相关性分析可知,对照组平均萌发时间与海拔呈正相关(r=0.847)。另外,随着染色体倍性的增加,其发芽势也随之变强,即倍性与发芽势呈正相关(r=0.541),这与赵宁等[18]的研究结果一致。发芽指数呈先高后低的趋势,且与海拔无显著性相关(p>0.05)。YY种子平均发芽时间显著高于另外两个倍性,说明作为云南山茶的原始倍性休眠状态较难打破,且与其他倍性种子差异显著(p<0.05)。ANOVA单因素分析显示,倍性间差异显著(F=6.53,p<0.05)。
由图1 B可见,种子的发芽率为ML>FM>YY,即经过相同时间低温层积处理的种子发芽率全部高于对照组,说明低温层积可以打破云南山茶种子休眠状态。且处理组与对照组发芽率差异显著(p<0.05),其中低温沙藏处理后的ML种子萌发率远高于另外两个倍性,即呈“低—高—低”萌发趋势,这与张梦颖等[17]的研究结果一致。随着染色体加倍,种子的萌发速度明显加快,这与赵宁等[18]的研究结果一致。随着海拔升高,种子的平均萌发时间逐渐缩短,即与倍性呈显著负相关(r=-0.84),这与屈燕等[15]的研究结果相同。对照组种子发芽势为ML>FM>YY,与海拔无显著相关性(p>0.05),而3个倍性云南山茶经过低温层积处理50 d后,发芽势均大于低温层积前的发芽势,说明低温层积可以显著提高不同倍性云南山茶种子萌发势。
2.3 不同浓度GA3对二倍体种子萌发特性的影响
GA3可以有效打破种子休眠,破坏阻碍萌发过程中的各种活性物质来提高萌发率、萌发势以及种子生活力等。由图2可见,在培养箱中恒温恒湿播种条件下,200 mg·L-1GA3处理后各项指标均高于其他浓度处理,这与邱鹏飞等[19]的研究结果一致。不同浓度GA3对二倍体种子发芽势的影响有显著性差异(p<0.05),这与李先民等[14]的研究结果一致。经过GA3处理的种子发芽率和发芽势显著高于对照组,0~400 mg·L-1范围内,随着GA3浓度的增加,二倍体云南山茶种子的发芽率与发芽势呈先升后降的趋势,这与张存虎[22]的研究结果一致。GA3在不同程度上显著提升了除发芽启动时间之外的各项指标。因此,GA3明显打破了云南山茶种子休眠。
2.4 海拔高度与低温层积对云南山茶种子萌发特性的双因素方差分析
如表3所示,低温层积对云南山茶不同倍性种子的萌发率以及发芽势的影响显著,但在平均萌发时间上无影响;另外海拔高度对种子萌发平均时间影响极显著,而对其他萌发指标无影响;双因素交互作用显示对所有萌发指标均有显著性影响。
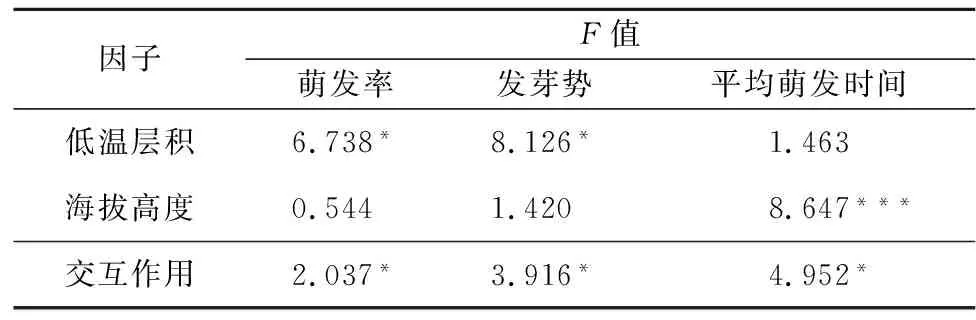
表3 海拔高度因子与低温层积的双因素方差分析
3 结论与讨论
3.1 倍性对云南山茶萌发特性的影响
杨继[20]认为,多倍体进化的结果一方面在很大程度上改善了不同基因组组分之间以及核基因组与细胞质基因组之间的相互协调关系,并提高了各倍性间相容性;另一方面也使得多倍体植物的适应进化、相同物种不同多倍体谱系的分化更加丰富多元。本研究结果表明,不同倍性云南山茶萌发特性及表型差异显著,云南山茶在多倍化进程中,并不是随着倍性增加其生物学特性增强,这与黄金艳等[21]的研究结果相同。这充分证明Stebbins[30]所指出的“每个物种都有一个最适的倍性,但绝大多数并非最高倍性”,也验证了顾志建[4]指出的“四倍体云南山茶分布范围、适应性较二倍体更广”的结论。因此推测四倍体植物内源激素更为复杂活跃。
种子表型性状是进行树木分类和遗传多样性研究的重要指标[22]。通过表型性状差异研究植物的遗传多样性报道较多,如对白皮松[24]、云杉[25]、蒙古栎[26]等的群体遗传多样性研究。本研究表明,处于不同居群的云南山茶各项表型性状及萌发指标间影响显著,随着染色体加倍,种子的含水量也随之增加,说明在长期演化过程中,为了适应恶劣的生存环境,云南山茶染色体发生变异加倍且表观形态也随之变化,形成多倍体云南山茶遍布我国西南地区乃至东南亚部分地区的局面。因此推测多倍化致使云南山茶抗逆能力较变异前增强。
3.2 不同前处理对云南山茶萌发特性的影响
鱼小军等[27]研究表明,低温层积可以很大程度上有效打破种子休眠机制,促进草本以及木本植物的种子萌发[27],但低温层积是否对同一物种的不同倍性产生不同影响,则因物种不同而有差异。罗弦等[28]研究发现,低温层积可显著提高4种苔草发芽率、发芽势等萌发指标,低温层积120 d可以有效打破苔草种子综合性休眠状态,从而达到萌发高峰期。李欣勇等[29]研究发现,低温层积两个月可显著提高球穗扁莎种子的各项萌发指标。本研究发现,低温层积50 d,对不同倍性云南山茶种子各项萌发指标均有明显提高,但对于各倍性的发芽启动时间均无显著影响。层积是否对不同倍性的发芽启动时间具有影响,有待进一步实验研究。
GA3能打破种子休眠,提高发芽率与发芽势,但对于激素处理的最适浓度、处理后种子平均发芽时间等问题,则因物种不同而存在差异。马艳华等[31]研究发现,白腊种子经30 ℃温水浸种12 h后再用80 mg·L-1GA3溶液浸种12 h,发芽率可达到68.3%。10~500 mg·L-1GA3可显著提前水青冈种子的萌发时间,其中100 mg·L-1溶液处理效果最好,发芽率为72.2%[32]。而经过10 mg·L-1GA3处理仙客来后的种子萌发指标与对照组萌发指标均存在显著差异(p<0.05)[33]。本研究结果表明,200 mg·L-1GA3浸种24 h可以使二倍体云南山茶种子的发芽率达到70%,且显著提高了发芽势并缩短了发芽时间,但随着浓度增加,种子的发芽启动时间越长,这可能是二倍体作为云南山茶的基础倍性,其内源激素复杂,GA3的介入打乱了种子正常萌发进程,具体原因有待进一步研究。
3.3 结 论
植物长期适应环境主要表现在表型及萌发特性差异形成多倍化现象,位于不同海拔生境居群已然形成不同倍性,不同倍性的云南山茶居群因长期存在于不同的生境,其所承受的生境压力也差异显著,最终会使各居群呈现出较显著的遗传多样性。导致群体间的遗传变异谱系化。但不同倍性云南山茶遗传多样性研究及各倍性种子的内源激素差异还需进一步研究。
