创伤后应激障碍小鼠海马神经细胞内FoxO3a和LC3蛋白的表达变化
2021-08-19王琼峰韩雨清包海军蒯锦霞蔡红星
王琼峰 韩雨清 周 阳 孙 卓 包海军 蒯锦霞 蔡红星
创伤后应激障碍(posttraumatic stress disorder,PTSD)又称延迟性心因性反应,是指突发性、威胁性、灾难性等刺激性生活事件或处境所导致个体延迟出现和长期持续存在的精神障碍。PTSD作为一种严重的精神疾病,其临床表现以闯入性创伤再体验症状(闪回)、回避和麻木类症状、持续性焦虑和警觉水平增高症状为主要特征[1,2]。
对PTSD患者的研究发现,在创伤刺激期间,海马等多个脑区在短期内可发生神经可塑性改变,神经细胞内钙离子调控紊乱,线粒体功能受损从而导致神经细胞受损[3]。在慢性刺激过程中,遭受刺激患者脑内自由基水平较正常增高,这会引起神经细胞脂质过氧化,导致由自由基介导的病理和随后的神经毒性,引起神经细胞凋亡、坏死、其他结构细胞损伤和修饰[4]。这些研究证明刺激反应会损伤大脑,引起脑内神经细胞的损伤。脑功能的预后与各种继发性损伤密切相关,如自噬等重要分子机制同样参与了脑神经细胞继发性死亡的过程。FoxO3a转录因子是Fox基因O亚家族的一个重要成员,其在细胞的增殖、凋亡、自噬、炎症等生物进程中发挥着重要作用[5]。近年来研究发现,FoxO3a是刺激反应中多个分子信号通路的交汇点,当机体遭受刺激时,FoxO3a能够通过影响自噬基因的表达来激活不同类型细胞的自噬机制。但目前在PTSD模型中,FoxO3a参与神经细胞自噬的调控较少有相关文献报道。本研究通过检测SPS刺激下小鼠的行为学改变及小鼠海马神经细胞内FoxO3a、LC3蛋白的表达情况,初步研究PTSD小鼠海马神经细胞自噬过程中FoxO3a蛋白的表达变化。
材料与方法
1.实验材料: (1)动物:成年雄性昆明小鼠68只,体质量25~30g,购自徐州医科大学实验动物中心,实验动物许可证号:SYXK(苏)2016-0028。(2)试剂:BCA蛋白含量检测试剂盒(编号KGP902)购自江苏凯基生物技术股份有限公司;超敏ECL化学发光试剂盒(编号P10100)购自苏州新赛美生物科技有限公司;兔抗Akt抗体(编号#4691)、兔抗p-Akt抗体(编号#13038)、兔抗FoxO3a抗体(编号#2497)、兔抗p-FoxO3a抗体(编号#9466)、兔抗LC3B抗体(编号#3868)购自美国Cell Signaling Technology公司;兔抗P62抗体(编号sc-25575)购自美国Santa Cruz Bio-technology公司;兔抗β-actin抗体(编号20536-1-AP)和山羊抗兔IgG二抗(编号SA00001-2)购自美国Proteintech Group公司。
2.动物分组:行为学检测(共20只),随机分为Control组和PTSD组,每组各10只;Western blot法检测实验(共36只),随机分为Control组(6只)和PTSD组,PTSD组依照1、3、7、14、28天5个时间点各设亚组,每个亚组各随机分配6只;免疫荧光检测实验(共12只),随机分为Control组和PTSD组,每组各6只。
3.PTSD动物模型的制备:首先小鼠适应性饲养1周,采用国际上公认的单一连续刺激(single prolonged stress,SPS)方法制备PTSD小鼠模型[6]:先将小鼠禁锢2h,之后立即将禁锢解除的小鼠强迫游泳20min(水深40cm,水温25℃),休息15min后进行乙醚麻醉至小鼠意识丧失,待其自然苏醒后分笼常规饲养,期间不再给予小鼠任何刺激。
4.旷场实验:本实验采用旷场实验箱,平均分为底面积相等的4个箱体,每个箱体规格50cm×50cm×40cm,箱体外周壁为黑色,靠壁底部区域面积组成外周活动区,其余部分区域面积组成中央活动区。测试前将小鼠放置于旷场实验箱中适应15s,实验开始时,将小鼠放置于旷场正中央,在操作者完成放开小鼠动作的同时,实验箱正上方的摄像机开始同步跟踪记录,记录时间为5min。本次旷场实验评估指标为小鼠的活动总路程(外周区域+中央区域)、中央区域活动的路程、中央区域停留的时间。
5.高架十字迷宫实验:高架十字迷宫装置由白色不透明有机材料制成,整个装置离地0.5m高,由两个开臂大小为长×宽(30cm×6cm)、两个闭臂大小为长×宽×高(30cm×6cm×15cm)及开臂和闭臂之间的中央区域(6cm×6cm)组成。实验开始时,将小鼠放置于高架十字迷宫的中央区域,头部统一朝向开臂方向,释放后实验人员迅速离开的同时,启动摄像系统开始跟踪记录,记录时间为5min,实验中跌落的小鼠则淘汰此数据。本次高架十字迷宫实验评估指标:小鼠进入开臂次数占总次数(进入开臂次数+进入闭臂次数)的百分比、小鼠在开臂停留时间占总停留时间(开臂停留时间+闭臂停留时间)的百分比。
6.Morris水迷宫实验:水迷宫为一个黑色圆形水池(直径105cm,高50cm),池内水深30cm,水温25℃左右,水池均等分为4个象限,将一个直径为6cm的白色透明圆形站台放置于靶象限(设为第一象限)内,站台位置距离池壁30cm,高度低于水面下1cm。具体实验步骤:(1)定位航行实验:每只小鼠每天训练4次,每次间隔1min,持续训练4天。实验前,去掉站台,让小鼠自由游泳60s以充分适应环境。实验开始时,将站台隐蔽于第一象限内,将小鼠头朝池壁随机从东北(NE)、东南(SE)、西南(SW)、西北(NW) 4个方向放入水池内,自由游泳60s寻找隐蔽的站台,60s内找到站台,记录所用时间为逃避潜伏期。若60s内未找到站台的小鼠,引导至站台,站台上停留30s,记录逃避潜伏期为60s。(2)空间探索实验:学习的第5天撤掉水下站台,从目标象限(站台)的对侧象限,将小鼠头朝池壁放入水池内,记录60s内小鼠的运动轨迹和目标象限(站台)停留情况。本次Morris水迷宫实验评估指标:60s内小鼠的逃避潜伏期、穿越原站台次数、原站台象限的停留时间。
7.Western blot法检测:冰台上操作剖离海马组织,预先按RIPA∶PMSF∶磷酸酶抑制剂=100∶1∶1的比例混合配制好蛋白组织裂解液,加入裂解液充分匀浆组织,冰上静置裂解30min,高速低温(13000r/min,20min)后取上清液。BCA法测定各组蛋白浓度,加入5×SDS上样缓冲液,95℃变性15min。配置SDS-PAGE凝胶,浓缩胶恒压80V电泳30min,分离胶恒压110V电泳至溴酚蓝到达分离胶底部,用电转法将蛋白质转移到PVDF膜上。5%脱脂奶粉溶液室温封闭2h;一抗4℃孵育过夜,TBST洗膜10min×3次;二抗室温孵育2h,TBST洗膜10min×3次。PVDF膜上滴加ECL化学发光液,暗室内曝光,凝胶成像分析系统成像。
8.免疫荧光染色实验:小鼠脑组织灌注取材,放置于4%多聚甲醛溶液中4℃固定36h,经20%、30%蔗糖溶液中4℃梯度沉糖脱水,冷冻切片机海马冠状切片,厚度20μm,-20℃保存备用。用时室温放置复温30min,PBS洗5min×3次;0.3%TritonX-100通透10min,PBS洗5min×3次;10%山羊血清封闭1h;一抗4℃孵育过夜,第2天室温孵育30min进行复温,PBS洗10min×3次;荧光二抗室温避光孵育2h,PBS洗10min×3次;DAPI复染核,室温孵育10min,PBS洗5min×3次;用含抗荧光淬灭剂的封片液封片,荧光显微镜下观察采集图像。

结 果
1.旷场实验检测:与Control组比较,PTSD组小鼠在旷场实验箱内活动总路程明显减少(P<0.01),其在中央区域活动路程、中央区域停留时间也均减少(P<0.01,图1)。
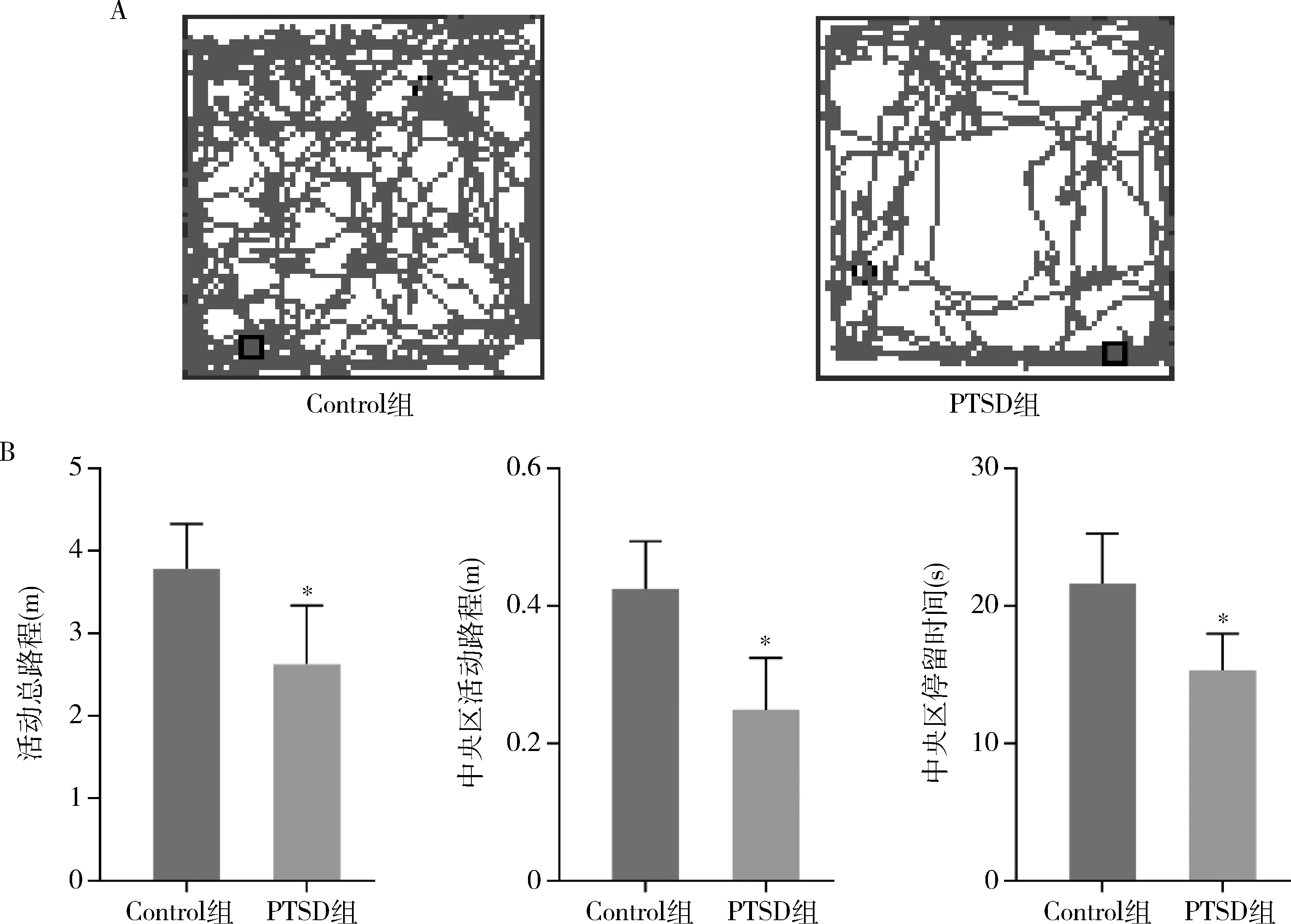
图1 两组小鼠旷场实验检测结果(n=10)A.运动轨迹结果图;B.两组小鼠在旷场箱内活动总路程、中央区域活动路程和停留时间的比较;与Control组比较,*P<0.01
2.高架十字迷宫实验检测:与Control组比较,PTSD组小鼠进入开臂次数百分比、开臂停留时间百分比均明显降低,两组结果比较差异有统计学意义(P<0.01,图2)。从小鼠的运动轨迹及测试分析结果发现,与Control组比较,PTSD组小鼠在闭臂内的活动总路程和总时间显著高于开臂。
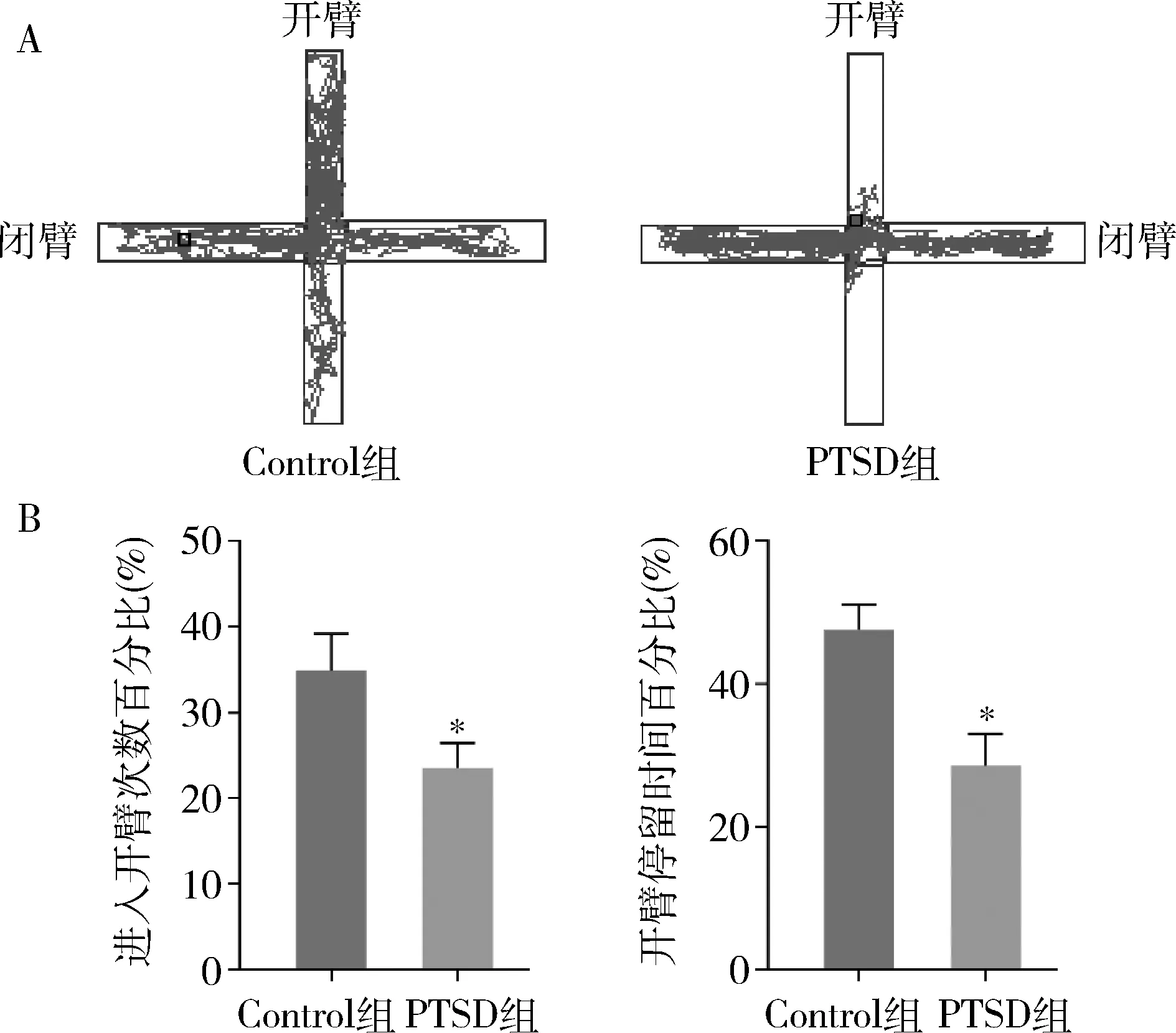
图2 两组小鼠高架十字迷宫检测结果(n=10)A.运动轨迹结果图;B.两组小鼠进入开臂次数和开臂停留时间百分比的比较;与Control组比较,*P<0.01
3.Morris水迷宫实验检测:在定位航行实验中,与Control组比较,PTSD组小鼠逃避潜伏期均明显延长(P<0.01);在空间探索实验中,与Control组比较,PTSD组小鼠穿越原站台的次数和原站台象限的停留时间明显减少(P<0.01,图3)。

图3 两组小鼠Morris水迷宫检测结果(n=10)A.运动轨迹结果图;B.两组小鼠逃避潜伏期、穿越原站台次数、原站台象限的停留时间比较;与Control组比较,*P<0.01
4.Western blot法检测自噬信号通路相关蛋白的表达:与Control组比较,PTSD组小鼠海马神经细胞p-Akt蛋白表达水平在1天时出现激活上调,之后表达开始下降;PTSD组海马组织FoxO3a蛋白表达增加,p-FoxO3a蛋白表达减少;PTSD组海马自噬相关蛋白LC3Ⅱ/LC3Ⅰ比值增加,p62蛋白表达水平降低,两组结果比较差异有统计学意义(P<0.05,图4)。SPS刺激后PTSD组各组蛋白表达随时间点变化而变化,FoxO3a蛋白能够在短暂SPS刺激下,迅速感受Akt蛋白的变化;SPS刺激后7、14天时间点,观察到PTSD小鼠海马LC3明显增多,FoxO3a蛋白表达水平也随之增加。

图4 Western blot法检测各组蛋白的表达情况(n=3)与Control组比较,*P<0.05,**P<0.01
5.免疫荧光检测小鼠海马神经细胞内FoxO3a和LC3的表达:荧光显微镜下观察发现,FoxO3a蛋白与CoraLite488标记的荧光二抗结合,其阳性反应呈现绿色荧光;LC3蛋白与CoraLite594标记的荧光二抗结合,其阳性反应呈现红色荧光;DAPI荧光染料对细胞核进行标记,发出蓝色荧光。在小鼠海马神经细胞内,FoxO3a和LC3蛋白呈现不同部位的表达分布,有的表达于胞质,有的可见表达于胞核;与Control组比较,PTSD组海马神经细胞内FoxO3a和LC3蛋白所呈现的绿色荧光和红色荧光信号强度均较Control组明显增强,且二者与DAPI叠加后的荧光强度也高于Control组(图5)。

图5 免疫荧光染色检测两组小鼠海马神经细胞内FoxO3a和LC3的表达情况(×400,n=6)A.海马神经细胞内FoxO3a的荧光表达;B.海马神经细胞内LC3的荧光表达
讨 论
PTSD疾病诊断难、发生率高、病程长、疗效差,是由刺激性事件引起的典型疾病之一,近年来成为了研究的热点,目前对于PTSD的治疗仅局限于心理、情绪方面的对症治疗,经治疗后患者的症状也很难完全消失,对于产生该问题的主要症结在于PTSD的发病机制尚未完全阐明。许多因素影响到PTSD的发生,如存在精神障碍的家族史与既往史、童年时代的心理创伤(如遭受性虐待、10岁前父母离异)、性格内向及有神经质倾向、创伤事件前后有其他负性生活事件、家境不好、躯体健康状态欠佳等,这些现象目前还在深入的研究中[7~9]。
有研究表明,PTSD患者的海马体积部分萎缩,提示PTSD发病过程中患者海马神经细胞发生了异常的损伤、死亡和丢失[10,11]。动物模型研究发现,海马部位存在有明显的神经细胞凋亡现象[12]。国内外研究者已经证实PTSD后神经细胞凋亡高峰并不发生在创伤刺激后的早期阶段,说明PTSD后神经细胞死亡为一长期、持续的继发性损伤过程,但本病的发病机制尚未完全阐明。PTSD发病过程中海马神经细胞的异常丢失和死亡除了与神经细胞凋亡有关外,是否同时存在其他损伤因素尚不明确。脑功能的预后与各种继发性损伤密切相关,神经细胞继发性损伤可造成脑组织结构与功能的进一步损害,是导致脑功能障碍的重要原因之一。反复发生的慢性刺激,可能通过一系列的病理生理变化、兴奋性毒性等因素引起随后的神经细胞死亡和神经功能紊乱,如运动、感觉、学习和记忆障碍[13,14]。
自噬现象是细胞内一种正常的生理活动,在维持中枢神经系统正常功能方面起着重要作用,在神经变性疾病中,自噬受损可能导致神经细胞的损伤[15]。在多数细胞正常情况下,自噬处于相当低的水平,但当机体处于刺激状态时,体内细胞的自噬可以被激活而呈现高水平,表现为自噬增强。当机体和体内的细胞遭受的刺激过多、程度过强时,自噬可以被过度激活,从而引起不可逆的细胞死亡,对机体造成损害,引起相关疾病的发生。
本研究通过SPS刺激方法建立PTSD小鼠模型,采用旷场实验、高架十字迷宫实验、Morris水迷宫实验检测SPS刺激后小鼠的行为学变化。旷场实验是检测动物焦虑水平的经典行为学方法,用来评定动物的警觉水平、焦虑状态和环境适应能力。本次检测结果发现,SPS刺激后小鼠的活动总路程明显减少,反映遭受刺激后小鼠对新环境的好奇感减弱,探索兴趣下降,同时,观察到小鼠常蜷缩在旷场箱体的角落旁,中央区域活动路程和停留时间减少,反映小鼠对开阔旷场环境保持警觉和害怕的心理。高架十字迷宫是检测动物焦虑和恐惧水平的实验设备,本次检测结果发现,SPS刺激后小鼠倾向于活动在两侧闭臂内,并喜好躲藏在闭臂角落处,反映遭受刺激后小鼠对重新体验创伤情景(高悬开臂危险)的回避,表现出恐惧水平较高,有明显的焦虑样行为。Morris水迷宫是检测动物空间学习和记忆能力的首选经典实验,逃避潜伏期是站台隐蔽试验中的重要检测指标,是反映小鼠寻找水下隐蔽站台所花费的时间,这个指标用时越短说明小鼠的空间学习和记忆能力越强,相反则说明小鼠的空间学习记忆能力受损。小鼠穿越原站台的次数和原站台目标象限的停留时间,这两个指标主要反映小鼠在较长时间内的记忆保持能力。本次检测结果发现,SPS刺激后小鼠找到站台的时间明显延长,运动轨迹紊乱,没有定向寻找站台的目的性,表明SPS刺激后小鼠的空间定位能力减弱,学习记忆能力受损。综合以上行为学结果可知,SPS刺激后小鼠表现出PTSD疾病的主要核心症状,即焦虑和恐惧水平提高,环境适应减弱,探索兴趣下降,逃避反应延迟,学习记忆能力下降。
近年来对FoxO3a的作用机制研究逐渐从肿瘤细胞转移到了神经细胞,FoxO3a在多种神经系统疾病中发挥着重要作用。大量研究发现,FoxO3a 在缺血引起的神经细胞凋亡中发挥重要作用,可作为治疗海马神经损伤及认知障碍的靶向基因[16]。在小鼠神经干细胞的研究中发现,FoxO3a基因还参与了促进神经细胞的分化[17]。在运动性神经疾病中,FoxO3a发挥神经保护作用,细胞核内FoxO3a的激活可以抑制由兴奋性神经毒素的表达引起的神经细胞死亡[18]。研究还发现,刺激引起的神经细胞损伤,其损伤程度在大脑不同细胞与区域间有差异,其中海马CA1区神经细胞尤其易受损伤,且随着年龄增长而加剧。Jackson等[19]研究认为,CA1区的易损性与FoxO3a在CA1区大量表达有关。
FoxO3a具有磷酸化和非磷酸化两种形式,其中非磷酸化形式的FoxO3a被认为具有转录激活活性分子,磷酸化在其活性及细胞内转移作用中起到关键作用,FoxO3a是PI3K/Akt信号通路下游的重要转录因子,在细胞应激研究中,当Akt磷酸化水平被抑制时,无法激活磷酸化的FoxO3a,使其与胞核输出蛋白14-3-3特异性结合受阻,导致FoxO3a在胞核内大量表达,从而增加细胞自噬[20]。同时,在本研究PTSD小鼠模型中,结果也发现,SPS刺激后小鼠海马神经细胞内p-Akt水平随时间延长表达下降,p-FoxO3a水平表达受阻,同时,观察到LC3和FoxO3a蛋白水平增加,荧光表达强度增强,结果提示PTSD疾病神经细胞自噬发生中可能存在FoxO3a的表达。但是由于PTSD后神经细胞自噬的具体作用机制相当复杂,FoxO3a与神经细胞自噬之间的关系仍有待于进一步研究。
