荔枝园荔枝蒂蛀虫常规爆发期的种群空间分布格局
2021-08-13全林发董易之李文景陈炳旭
全林发,董易之,徐 淑,李文景,陈炳旭
(广东省农业科学院植物保护研究所,广州 510640)
荔枝在我国具有悠久的栽植和食用历史。建国以来我国荔枝种植面积从6 667 hm2扩展到551 746 hm2,增长了82.7倍,产量和产值都得到了跨越式增长,基本形成了海南特早熟优势区、粤西中晚熟优势区、粤中桂东南中晚熟优势区、粤东闽南晚熟优势区、长江上游特晚熟优势区和云南高原立体生产优势区六大产区(陈厚彬和齐文娥, 2019)。荔枝蒂蛀虫ConopomorphasinensisBradley属鳞翅目Lepidoptera细蛾科Gracillariidae,以幼虫钻蛀为害荔枝、龙眼的果实、花穗、嫩梢和嫩叶,尤其偏爱为害果实,一年发生9~12代,世代重叠严重(Yaoetal., 2018)。随着荔枝、龙眼产业在中国的蓬勃发展,荔枝蒂蛀虫已经取代荔枝蝽成为荔枝龙眼园内的第一大害虫(李文景等, 2018)。一般年份被荔枝蒂蛀虫为害的荔枝、龙眼虫果率为10%~20%,严重时达60%~90%,使荔枝龙眼产业蒙受巨大损失(李康, 2002; 陈炳旭等, 2011)。
空间格局是昆虫种群的重要特征之一,它是由昆虫种群的生物学特性与特定生境条件相互作用协同进化的结果。研究昆虫的空间格局不仅可以揭示种群内和种群间的空间结构,而且还可以为抽样设计、种群消长趋势和扩散范围的预测预报以及制订害虫防治策略提供较好的依据(宗世祥等, 2006)。随着空间生态学理论和地理统计学方法的发展,害虫种群的空间分布格局及种群扩散趋势能够得到精确的解析和计算(丁岩钦, 1994; 宗世祥等, 2005)。地理统计学是在地质分析和统计分析互相结合的基础上形成的一套分析空间相关变量的理论和方法,能最大限度地利用田间调查所提供的各种信息及空间数据,相比经典的统计学方法更为精确,且能有效避免系统误差(Liebholdetal., 1993; 周强等, 1998)。通过直接测定和分析变量而建立起的预测模型,能有效解决昆虫种群的空间异质性问题(Naranjo和Flint, 1995; 李建庆等, 2018)。目前,荔枝蒂蛀虫的研究工作主要集中在生物学特性及防治技术上,研究不深且涉及的领域较窄,有关荔枝蒂蛀虫种群空间分布格局的研究尚未有报道(李文景等, 2018)。笔者对广东省广州市天河区华南农业大学荔枝栽培试验基地荔枝园荔枝蒂蛀虫卵、蛹和成虫虫口密度及发生特点进行了调查,采用生物学统计方法和地理统计学方法对荔枝园中荔枝蒂蛀虫卵、蛹和成虫常规爆发期的种群空间分布格局进行了较全面的研究,为了解荔枝蒂蛀虫在荔枝园中的空间分布特征,提高抽样技术及种群的监测与防控提供理论指导。
1 材料与方法
1.1 样地设置与调查方法
1.1.1样地基本情况
试验地位于广东省广州市天河区华南农业大学荔枝栽培试验基地(E113°21′, N23°09′),该地区属于南亚热带海洋季风气候,光热资源充沛,年平均气温在21.9~22.8℃之间,霜日少。降水充足,年平均降水量在1 384.4~2 278.3 mm之间。调查样地为人工栽植的荔枝园,面积约为120 m×70 m,主栽荔枝中晚熟品种,包括桂味、糯米糍等。荔枝树平均树龄为27年,平均树高3.2 m。为恢复调查样地的原生态性,保证荔枝蒂蛀虫田间自然种群的稳定性,本试验调查前三个月内未使用任何杀虫剂。
1.1.2调查方法
由于该果园排列规律,采用坐标法逐株系统调查,把果园中的200多棵果树按20列×10行网格法矩形排列,以左下角第1棵树为坐标原点(0,0),以北向和东向建立直角坐标系,于坐标纸上将坐标系划分为同样大小的各个小栅格,将调查荔枝树的位置标记在坐标纸上相对应的小栅格内,每个小栅格代表1个样本的实际观测值。依据荔枝蒂蛀虫的生物学特性及其田间预测预报结果,在虫害发生的高峰期展开相关调查。
为了解中晚熟荔枝品种荔枝蒂蛀虫危害盛期种群的空间分布格局,分别于2019年5月8至12日和6月4至8日,在荔枝蒂蛀虫成虫发生高峰期,对坐标原点为 (5,2)、(5,6)、(16,2)、(16,6)四个样点处12列×5行共60棵桂味和糯米糍荔枝树进行逐株系统调查3次,每次调查间隔1 d。每棵树按照东、南、西、北、中5个方位,每个方位固定2个主枝(即着生于中心骨干枝上的的第2级骨干枝),用目测法分别调查荔枝蒂蛀虫成虫和卵的数量;成虫调查时采用目标枝条搜寻法,必要时敲击或轻微摇晃目标枝条,以枝为单位记录静栖及飞舞的荔枝蒂蛀虫成虫的数量,并统计有成虫分布的荔枝树株数,计算样方内荔枝树有虫株率及含不同成虫密度的荔枝树占比;卵调查时在每个目标果枝上随机选取10颗荔枝果实,采用手持双目镜调查并记录果实表面的着卵粒数,将各组的计数结果综合在一起,计算每棵树荔枝蒂蛀虫卵的平均密度。又分别于2019年5月21-25日和6月16-20日,荔枝蒂蛀虫蛹期内,在相同的样方内,应用调查成虫和卵的方法,以枝为单位,逐枝、逐叶记录整个枝条上蛹的数量(刘军和, 2009)。
另外,在上述矩形样方内,于2019年5月8日和6月8日,采用坐标法逐株系统调查样方内荔枝树的挂果情况。调查方法为:每棵树按照东、南、西、北、中5个方位,每个方位随机选择4个结果枝(即果树第3~4级枝),共调查20个枝条的挂果情况。以正常挂果的枝条作为挂果枝,统计每棵树挂果枝条的数量,求得每棵树的挂果率,计算公式为:果树挂果率(%)=挂果枝条数/调查总枝条数×100。
1.2 空间格局的地统计学分析
当一个变量在空间上与其位置有关时称为区域化变量,区域化变量在空间上因其相互间的位置关系或相关性而存在一定的规律性变化称为空间变异(王其武, 2016)。半变异函数是研究区域化变量的主要工具,被定义为变量Z(xi)和Z(xi+h)增量平方的数学期。其计算公式为:γ(h)=1/2N(h)∑[Z(xi)-Z(xi+1)]2,式中,i=1,2,…,N(h),γ(h)为相对距离的半变异函数值,N(h)是被h分割的数据对(xi,xi+h)的对数,Z(xi)和Z(xi+1)分别是点xi和xi+h处样本的测量值,h为两样点间的距离(李建庆等, 2016)。本研究中的变量值分别为荔枝蒂蛀虫卵(粒/株)、蛹(个/株)和成虫(头/株)的密度。
半变异函数理论模型的拟合:半方差γ(h)是距离h的函数,以距离h为横坐标、半方差γ(h)为纵坐标构成的二维平面图称为半变异函数图。半变异函数曲线图的形状能反映变量空间分布的结构及相关类型,以及空间相关范围的大小,一般常用的模型有表征所研究种群为聚集分布的球型、指数型、高斯型,以及表征种群呈随机分布或均匀分布的直线型(胡瑜珩, 2015)。前者指半变异函数值未达到稳定状态之前,空间依赖性的大小随采样距离增大而逐渐降低;后者表明在抽样尺度下没有空间相关性(汪文俊等, 2012; 胡瑜珩, 2015)。具体选择采用哪种理论模型主要依据决定系数(R2)、残差平方和(RSS)、变程(A)和块金系数(C0)的大小依次判断,从而得到最优拟合模型(孙鹏举等, 2015; 刘永华等, 2018)。其中块金系数(C0)大小反映了变量的随机程度;基台值(C+C0)大小可反映变量变化幅度的大小;空间结构比率[C/(C+C0)]反映空间结构系数在样点变量差异中所占的比率,以此值大小可判定系统内变量的空间相关程度:介于25%~75%之间为中等程度的空间相关性,大于75%则具有较强的空间相关性(李建庆等, 2016)。
空间数据差值:空间分布的区域化变量用Kriging插值法生成分布图,可以反映调查样点之间区域化变量的分布状况。利用Surfer软件中Grid功能、Kriging法对荔枝蒂蛀虫空间分布的区域化变量进行插值分析(孙鹏举等, 2015; 胡瑜珩等, 2015),通过Surfer 12软件生成空间分布等值线与矢量叠加图(刘永华等, 2018)。
1.3 聚集成因分析

1.4 数据统计
应用Excel 2007对基础数据进行统计分析,依据相关统计学理论和方法将调查所得数据进行归类整理。利用GS+9.0软件实现半变异函数理论模型的拟合;采用Surfer软件中Grid功能、Kriging法对害虫种群空间分布的区域化变量进行插值分析,并通过Surfer 12软件生成空间分布等值线与矢量叠加图。
2 结果与分析
2.1 不同时期荔枝蒂蛀虫成虫的密度分布
5月份和6月份调查样地中挂果/无挂果荔枝树荔枝蒂蛀虫成虫的密度分布情况见图1和图2,结果显示:5月,无挂果树和挂果树的有成虫株率分别为67.74%和91.67%,其中成虫密度>5头/株的样本比例分别为16.13%和36.11%;6月,无挂果树和挂果树的有成虫株率分别为68.57%和90.91%,其中成虫密度>5头/株的样本比例分别为8.57%和9.09%。比较挂果树和无挂果树上荔枝蒂蛀虫成虫密度发现,挂果树成虫密度明显高于无挂果树(见图1、图2)。
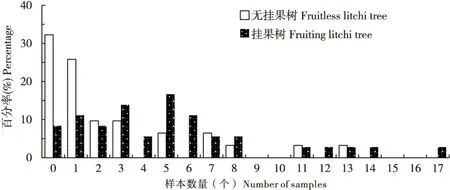
图1 5月份样方内荔枝蒂蛀虫成虫密度分布直方图Fig.1 Histogram of density for the Conopomorpha sinensis adults in quadrat in May

图2 6月份样方内荔枝蒂蛀虫成虫密度分布直方图Fig.2 Histogram of density for the Conopomorpha sinensis adults in quadrat in June
2.2 荔枝蒂蛀虫不同虫态的空间分布特征
采用种群密度等值线图可以直观显示荔枝蒂蛀虫发生程度在样地内的空间分布特征,在样地中对坐标原点为(5,2)、(5,6)、(16,2)、(16,6)4个样点处12列×5行共60棵树调查的数据绘制等值线与矢量叠加图,结果如图3所示。从5月中旬至6月中旬,样方内荔枝树的挂果率有所下降,而荔枝蒂蛀虫不同虫态的空间分布格局也发生相应的变化。总体上,样方内荔枝蒂蛀虫卵、蛹和成虫均呈现出不同程度的聚集分布特点:样方内有1个或多个聚集斑块,每个斑块的虫口密度由中心区域向四周逐渐减少,反映了该虫在样地内的聚集强度、空间尺度和扩散方向。与成虫相比,荔枝蒂蛀虫卵和蛹的聚集斑块少且集中,聚集分布趋势更强,成虫的聚集斑块较多且分散,聚集程度不太强。另外,对比荔枝树挂果率和虫害密度的等值线图发现,荔枝蒂蛀虫卵和蛹的聚集斑块中心与荔枝树高挂果率聚集中心大同小异,可见,该虫的田间空间分布与荔枝树挂果情况密切相关。

图3 荔枝蒂蛀虫不同虫态的等值线图与矢量叠加图Fig.3 Overlay maps of isoline and vector for Conopomorpha sinensis at different stages
2.3 荔枝蒂蛀虫不同虫态的空间分布型分析
荔枝园调查数据的基本统计参数见表1。利用GS+9.0软件对荔枝蒂蛀虫卵、蛹和成虫的空间分布情况进行处理,选取R2最大、RSS最小的变异函数理论模型作为最佳拟合模型,各拟合参数及分布型见表2。结果表明:5月份,调查样地内荔枝蒂蛀虫卵、蛹和成虫的空间分布有明显空间结构,拟合模型分别球型模型、球型模型和指数模型,即3种虫态的空间分布型均为聚集分布。卵、蛹和成虫的半变异函数决定系数R2分别为0.948、0.971和0.766,各模型的拟合程度较高。卵、蛹和成虫的变程在5.63~37.41 m之间,空间依赖范围未超出研究尺度,且在此距离范围内调查样地中任何两点卵、蛹和成虫的数量都各自存在一定的相关关系,空间相关性随着距离的增大而逐渐减小,空间结构比率在71.28%~91.53%之间,均值为78.37%,大于75%,表明该月份荔枝蒂蛀虫在荔枝园内具有较强的空间相关性,具体到各虫态,卵和蛹均大于75%,空间相关性较强,成虫介于25%~75%之间,为中等程度的空间相关性。

表1 荔枝园调查数据的基本统计参数

表2 荔枝蒂蛀虫不同虫态的变异函数理论模型、拟合参数及空间格局类型
相似的,6月份调查样地内荔枝蒂蛀虫卵、蛹和成虫的空间分布型均为聚集分布,拟合模型分别为指数模型、球型模型和指数模型,R2介于0.615~0.825之间,拟合程度较好。卵、蛹和成虫的变程在5.28~51.81 m之间,空间依赖范围未超出研究尺度,调查间隔距离均在其变程范围内,明显小于其变程,空间依赖型明显,空间结构比率在50.03%~60.93%之间,均值为54.77%,介于25%~75%之间,表明该月份调查样地具中等程度的空间相关性,具体到各虫态,卵、蛹和成虫均介于25%~75%之间,为中等程度的空间相关性。
2.4 荔枝蒂蛀虫在荔枝树上聚集成因分析

表3 荔枝蒂蛀虫不同虫态的聚集原因分析
度的下降,样地内聚集分布的成虫的λ值为1.48<2,由此可见该时期成虫的聚集仅由环境条件引起。
3 结论与讨论
空间格局是昆虫种群的重要属性之一,由物种的生物学特性和环境条件所决定,其反映了昆虫在寄主植物和非生物环境影响下,空间需求的内在生物学特性(李哲等, 2004; 鄢铮和王正荣, 2011)。了解昆虫的空间分布及动态变化对有效的进行虫害管理与防治具有重要意义(孙鹏举等, 2015)。而昆虫种群的空间分布型不仅与昆虫自身密度、扩散活动能力、生活习性有关,而且与寄主物候期、栽培特点及其生长的地理环境有着密切的联系(贾楠等, 2012;胡瑜珩等, 2015;李建庆等, 2018)。
田间调查发现,5月至6月,正值珠三角地区中晚熟荔枝品种果实膨大期至果实着色初期,是荔枝蒂蛀虫种群常规危害盛期及防控的关键期。为此深入掌握该危害阶段荔枝蒂蛀虫的种群数量动态和空间分布格局,对于揭示害虫的危害特性与发生发展规律,采取有效防治措施及精准施策控制虫害扩大化具有非常重要的意义。荔枝蒂蛀虫以幼虫蛀果为害,隐蔽性强,其危害的寄主树体较大,单株受害树虫口数量多,涉及相关种群数量调查时,多数以单株树木作为调查单位,单株树木即可看作是一个调查样方(李建庆等, 2018)。此外,对地理统计学而言,每个样地30个样方即可达到数据统计要求,本研究设调查样本为60棵树,针对不同虫态发生高峰期动态重复调查3次,综合考虑调查成本和统计需要,能够反映调查种群数量统计需要。本研究首次利用地统计学方法对荔枝蒂蛀虫卵、蛹和成虫在荔枝品种园内的空间分布进行了分析,结果表明该虫3种虫态在荔枝园内的均呈现较明显的聚集分布,同时发现荔枝树挂果率对其卵、蛹和成虫在聚集上有很大的影响,这符合了荔枝蒂蛀虫的相关生物学特性。通过聚集指数λ分析也表明,荔枝蒂蛀虫卵和蛹在荔枝园的聚集的原因不仅受环境条件影响,还与自身生物学特性密切相关。
昆虫种群的空间分布状况与种群的扩散活动能力有着直接关系(曾凡勇等, 2012)。荔枝蒂蛀虫主要以幼虫蛀食果实为害,田间成虫多在荔枝果实而较少在叶片上产卵(刘少兰, 2012; 董易之等, 2018)。由于荔枝蒂蛀虫卵本身没有迁移能力,所以荔枝蒂蛀虫卵空间格局主要依赖成虫的产卵场所,因此卵的空间格局与成虫的空间格局相近。另外,田间调查发现,荔枝蒂蛀虫成虫多产卵于果壳基部(果蒂),这样有利于幼虫孵化后钻入果实为害,从而造成挂果树成虫密度明显高于无挂果树,这也是引起荔枝蒂蛀虫成虫在挂果荔枝树上的聚集分布趋势的环境因素之一。而荔枝蒂蛀虫卵孵化后随即钻蛀进入植物组织中,至老熟化蛹前始终在寄主植物内部取食,幼虫老熟后咬穿植物爬出,就近叶片正面中脉近叶尖处或地面的枯枝落叶处结茧化蛹,这些都可能是荔枝蒂蛀虫蛹呈聚集分布的一个生物学原因(陈炳旭等, 2011)。但是,本次调查发现,5月荔枝园内成虫虫口密度较高时,园内成虫聚集的原因主要是由于成虫自身生物学特性(成虫喜阴怕光、产卵选择行为等)及生境共同作用所引起的;但6月随着成虫虫口密度下降,荔枝园内成虫的聚集分布主要由生境引起。笔者推测,荔枝蒂蛀虫趋向于以挂果荔枝树为中心的聚集分布是“荔枝蒂蛀虫-寄主”高度协同进化的结果,同时反映了昆虫对植物的利用策略(钦俊德, 1987; 陆宴辉等, 2008)。为此,通过可视化分析害虫聚集点及扩散方向可以有效指导害虫防治。另外,该虫的种群发生动态和空间分布格局同时受寄主植物的物候期、树势、果园郁闭程度、光照、温度、湿度及天敌昆虫等因素影响。
对种群分布格局进行可视化的定性分析是种群分布格局研究中常用的手段之一,并能形象描述害虫在特定空间结构上的分布状态,对更好地研究害虫的防治方法具有一定的指导意义(付园园等, 2019)。荔枝蒂蛀虫是我国最重要的荔枝、龙眼蛀果类害虫之一,该虫在珠江三角洲中晚熟荔枝优势区,每年发生10~11代,成虫第一代始发于3月中旬前后,第3代开始世代重叠,5~7月发生危害最重,多以幼虫在荔枝冬梢或早熟品种花穗顶端轴内越冬(陈炳旭等, 2017)。本研究应用种群分布特点及空间分布信息,针对荔枝蒂蛀虫常规爆发期(5~6月),阐明了该虫卵、蛹和成虫在荔枝园中空间分布格局,通过Kriging法获知害虫聚集点及扩散方向以便对园区内荔枝蒂蛀虫种群进行有效监测和防控。由于时间和其它条件的限制,本文仅对5月至6月中晚熟荔枝园内荔枝蒂蛀虫的空间分布型进行了研究,而不同品种、栽植模式果园、单一品种园与品种混种园等的虫害发生和种群空间布局是否存在差异,对于不同虫态在果园内的最佳抽样技术该如何确定,值得进一步研究。另外,应用地理统计学对荔枝蒂蛀虫种群在时间上的连续性与依赖性的研究今后还需增加调查次数,以获得更多资料进一步完善。
综上所述,5月至6月底,荔枝蒂蛀虫卵、蛹和成虫在荔枝园内趋向以挂果荔枝树为中心的聚集分布,聚集斑块由聚集中心区域向四周逐渐减小。该时期荔枝蒂蛀虫卵和蛹的聚集均是由生境和自身生物学特性(幼虫蛀果为害和成虫产卵选择性等)共同引起的,但成虫在虫口密度降低时,其聚集分布仅由生境引起。
