椰子织蛾触角感器的超微结构观察
2021-08-13刘青钊阎雄飞马光昌
刘 丽,刘青钊,阎雄飞,马光昌,刘 博,杨 帆,阎 伟*
(1.中国热带农业科学院椰子研究所,海南文昌 571339;2. 榆林学院生命科学院,陕西榆林 719000;3. 中国热带农业科学院环境与植物保护研究所,海口 571101;4. 海南大学林学院,海口 570228)
椰子织蛾OpisinaarenosellaWalker,又名椰子木蛾、黑头虫,属于鳞翅目Lepidoptera木蛾科Xyloryctidae,是椰子Cocosnucifera、海枣Phoenixdactylifera等棕榈科植物上重要的检疫性食叶害虫(李后魂等, 2014; 赵力等, 2015)。该虫原产于亚洲的斯里兰卡和印度,现主要分布于东南亚国家和地区。可危害椰子等34种棕榈科植物,此外还可危害香蕉Musanana、甘蔗Sacharumofficinarum等重要农林经济作物(阎伟等, 2013; 吕宝乾等, 2013)。椰子织蛾以幼虫危害寄主,使受害寄主叶片卷折、枯萎焦黄,形状呈现火烧状。危害严重时将整株椰子树叶片取食干净,致使树势减弱,落果量增加,产量减少(Murthyetal., 1995; Muralimonan and Srinivasa, 2005; 方萍等, 2018)。该虫适应能力强,传播快,自2013年8月首次在海南省发现该虫入侵危害以来,现已在我国海南省大部分地区、广东省和广西省等地迅速定植蔓延危害(阎伟等, 2013)。如果不加强椰子织蛾成灾机理和控制技术研究,将给我国棕榈科植物种植区域造成不可弥补的经济损失、景观损失和生态损失,严重影响棕榈科植物相关产业的发展(吕宝乾等, 2013)。
触角在昆虫寻找寄主、取食产卵、寻找配偶、交尾、感受环境刺激过程中起着重要作用(Schneider, 1964; 马瑞燕等, 2000)。触角上感器具有感受物理刺激(温度、湿度、风速)和化学刺激(植物挥发出的信息化合物和昆虫自身挥发出的各种信息素)的功能(尹文英和郦一平, 1980; 马瑞燕等, 2000)。近年来,利用昆虫触角的感器感受化学刺激,来探索昆虫行为产生的机理,已经成为害虫可持续控制的新途径和研究的国际热点。鉴于此,明确椰子织蛾触角感器的超微结构、类型和分布情况,有利于明确椰子织蛾寄主选择和寻找配偶的化学通讯机理,在此基础上,进一步发展该虫的行为控制技术,对椰子织蛾的科学可持续防控有重要意义。
随着科学技术的高速发展,触角感器逐渐成为研究热点,很多科学家应用扫描电子显微镜技术、透射电镜技术(Transmission Electron Microscope,TEM)、免疫组化定位、转录组测序等技术对触角感器的超微结构、生理结构、感受功能等方面进行了大量的研究(郭振营和王振军, 2015)。鳞翅目是昆虫纲中第2大目,种类繁多,很多为农林生产中的重大害虫。当前很多研究关注于鳞翅目昆虫触角感器的研究,已涉及29科60多种鳞翅目昆虫,例如棉铃虫Helicoverpaarmigera(尹文英和郦一平, 1980)、桃小食心虫Carposinasasakii(宫田睿等, 2012)、小地老虎Agrotisypsilon(向玉勇等, 2016)、舞毒蛾Lymantriadispar(马菲等, 2013)、栎黄枯叶蛾Trabalavishnou(靳泽荣等, 2016)等。张国辉(2012)报道梨小食心虫Grapholithamolesta触角上分布有7种类型感器,雌虫和雄虫无明显差异。郭永福等(2018)按照新的分类系统对梨小食心虫Grapholithamolesta感器重新进行了研究,发现该虫有8种类型感器,耳形感器数量在雌成虫和雄虫上分布不同,雌雄虫的刺形感器、毛形感器等6种感器的长度存在显著差异。陈丹丹等(2017)对核桃举肢蛾Atrijuglanshetaohei触角上感器进行了系统的研究,发现该虫触角上具有9种类型的感器。龚粤宁等(2018)报道荔枝异形小卷蛾Cryptophlebiaombrodelta触角上有7种感器,毛形感器是主要感器。
椰子织蛾是近年入侵我国的重要检疫性以叶片为食的害虫。该虫主要危害椰子等棕榈科植物,取食后会形成丝状虫道,危害2~3 d后,致使叶片自动向下卷折闭合,为该虫提供了隐蔽的场所,使化学农药无法接触,鸟类等捕食性天敌难以发现,造成该虫防治非常困难的现象。很多科学家对其形态特征、适生区、生物防治、化学防治等方面进行了研究报道(Andoetal., 1981; Cock and Peprera, 1987; Sujatha and Singh, 2004; Venkatesanetal., 2009; 李洪等, 2015; 阎伟等, 2015)。关于椰子织蛾感器方面尚未见报道,鉴于此,本研究以棕榈科植物检疫性害虫椰子织蛾为研究对象,对该虫触角进行光学显微镜和扫描电镜观察,系统研究了触角的外部形态、感器类型和分布,为揭示该虫化学通讯机制提供基础资料,从而为该虫的防控措施提供理论依据。
1 材料与方法
1.1 供试虫源
椰子织蛾蛹来自中国热带农业科学院椰子研究所和中国热带农业科学院环境与植物保护研究所。原始种群采集自不同时期海南椰子织蛾分布的7个市县的多种寄主植物。将蛹置于人工气候箱温度为26~28℃,相对湿度为45%~65%中养虫笼内羽化,收集刚羽化的椰子织蛾雌雄成虫供试。
1.2 供试药品及用具
所需药品:氢氧化钠分析纯(天津奥普升化工有限公司)、95%乙醇和无水乙醇为分析纯(天津市富宇精细化工有限公司)
所需用具:导电胶(顺生电子科技有限公司),手术刀(深圳美鑫电子公司)、样品台(顺生电子科技有限公司)、养虫笼(河南智科弘润仪器有限公司)
1.3 供试仪器
椰子织蛾饲养和触角观察所需要的仪器、型号和生产商见表1。
表1 试验所需要主要仪器、型号和生产商
1.4 椰子织蛾触角的制备
光学显微镜样品制备:将椰子织蛾成虫置于培养皿中,在0℃下冷冻5~8 min,置于XTB-I体视显微镜下,将椰子织蛾的头部以上整体切下,用解剖镜直接观察触角各节的颜色。由于触角上覆盖着鳞片,节与节之间不清晰,触角感器各节用氢氧化钠处理后进行测量。具体步骤为将切好的触角用蒸馏水清洗2~3次后,放在13%氢氧化钠浸泡6 h,用蒸馏水洗净后备用。
扫描电镜的样品制备:椰子织蛾成虫触角的切除同光学显微镜样品的处理,放置于50 mL的烧杯中,用流动的蒸馏水清洗3~4次,再用超声波清洗6~8 min。之后参考郭永福等(2018)对梨小食心虫的方法,用乙醇和丙酮进行逐级梯度脱水,分别脱水12 min,最后用丙酮脱水2次,置于无尘玻璃罩中自然条件下干燥,用牙签将5根触角的背面、腹面分别固定在粘有导电胶的载物台上备用。
1.5 椰子织蛾触角的光学显微镜观察
触角颜色的观察:将处理过的触角干燥后,直接放置于Nikon SMZ 18光学显微镜下,观察触角的外部形态及颜色。
触角各节的长度测量,用氢氧化钠处理后的样品,置于显微镜下,测量触角柄节、梗节和鞭节各节的长度,并拍照记录。
1.6 椰子织蛾触角扫描电镜观察
将之前处理好的载物台置于TY2016000397离子溅射仪上,进行喷金染色,将载物台旋转,力求触角各个角度都均匀镀金(靳泽荣等, 2016)。然后使用HITACHI S-4300N扫描电子显微镜,调节电压为5 000 v,放大210~75 000倍观察,对椰子织蛾感器进行观察和拍照。
1.7 图像处理
图象处理:所有图像都使用Photoshop CS 6.0和Microsoft Office PowerPoint 2016进行处理,触角各种类型感器长度、基部直径和感器窝的直径的平均值和标准误差,用DPS 7.05软件进行分析处理。比较方法采用Ducan法。
2 结果与分析
2.1 椰子织蛾雌、雄成虫触角形态特征
椰子织蛾雌雄成虫触角均呈线状,上面覆盖着鳞片,长度为4.8~7.2 mm,由柄节(Sc)、梗节(Pe)和鞭节(Fl)构成,鞭节由36~40亚节组成(表2)。柄节呈灰黄色,雌虫和雄虫触角柄节平均长度分别为0.43±0.06 mm和0.48±0.05 mm;梗节比柄节略短呈现灰褐色,雌虫和雄虫触角梗节平均长度分别为0.38±0.07 mm和0.42 mm,鞭节平均总长度为7.10±0.06 mm和7.34±0.04 mm,灰色和灰褐色相间,亚节间镶嵌着黄绿的条形斑点;前15节黑白相间,剩余节呈淡绿色,中间各节呈圆柱形,最后一节呈锥形。
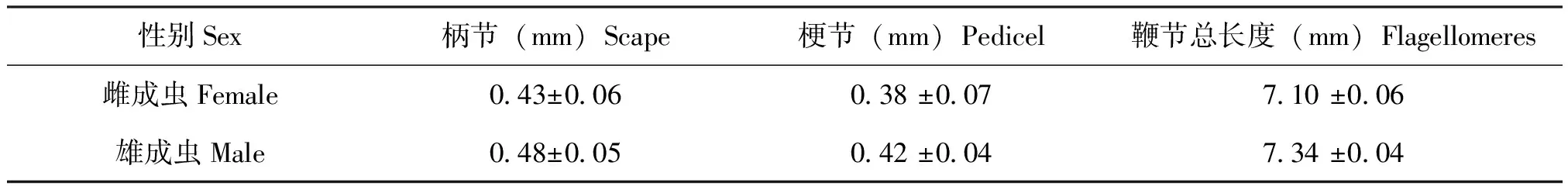
表2 椰子织蛾成虫触角各节的平均长度
鳞片是椰子织蛾触角覆盖最多的附生物,除柄节基部和鞭节最后亚节端部外,覆盖着整根触角表面,背面分布数量明显高于腹面,基部有短柄着生在基窝内,端部呈波浪状或弧形。鳞片表面有较深的纵脊,未发现有小孔(图1-A)。根据鳞片的长度和宽度不同,可以分为两种类型:长型鳞片(图1-B)和短型鳞片(图1-C)。长型鳞片,长度为26.7~56.6 μm,中部宽度为12.0~15.22 μm,基窝宽度:1.1~3 μm,主要分布在柄节的中部,鞭节最后亚节的末端。短型鳞片,长度为70.1~106.3 μm,中部宽度为10.87~14.3 μm,基窝宽度:1.52~3.3 μm。主要分布在柄节上部、鞭节各亚节的中部。

图1 椰子织蛾成虫触角鳞片Fig.1 The scales of antennae of Opisina arenosella注:A,触角表面鳞片分布;B,长形鳞片;C,短形鳞片。Note: A,Distribution of scales on antennal; B, Elongated scales; C, Short scales.
2.2 椰子织蛾触角感器的类型、分布及形态特征
(1)刺形感器(Sensilla chaetica)
刺形感器的形状像刺一样,明显高于周围其它感器。感器表面有环状纹理、不规则浅沟或光滑,感器壁无任何孔,根据长度和与触角表面的夹角可以分为刺形感器I和刺形感器II。
刺形感器I,明显高于其它类型感器,大部分感器与触角表面呈40°~75°角,基部有一凸出的感器窝为臼状,末端钝尖,鞭节末端亚节最为明显。感器表面有较浅的环状螺纹,接近基部逐渐消失,基部有不规则浅沟,感器壁无气孔。感受器的平均长度为32.8~57.27 μm,基部宽度为2.73~4.17 μm,感器窝为臼状,宽度为3.33~4.36 μm。主要分布于触角鞭节各亚节的腹面中部、侧面和鞭节最后亚节接近端部。在鞭节背面数量较少,通常为4~6根,分布于鞭节各亚节连接处和鳞形感器周围(图2-A)。
刺形感器II,感器直立或几乎直立于触角表面,感器与触角表面呈80°~90°角,感器窝为浅的臼状窝,感器末端急尖,感器表面光滑或具有浅的环状纹,感器壁无孔。感受器的长度为12.4~26.8 μm,基部宽度为1.1~2.7 μm,主要分布于柄节的基部侧面,雄虫比雌虫略长(图2-B)。
(2)毛形感器(Sensilla trichodea)
毛形感器的形状像单根毛发一样,端部卷曲,感器基部有明显感器窝,感器表面有较深的纵纹或者竖纹,感器顶端和壁布满小孔。按照感器长度不同和形状差异不同可以分为:毛形感器I和毛形感器II。
毛形感器I,在椰子织蛾触角上分布数量较多,通常和刺形感器混生在一起,与触角表面的夹角小于32°,感器端部钩状,中部表面有环状竖纹,接近基部有纵纹,感器基部有明显的感器窝。感受器的平均长度为27.4~46.53 μm(雌虫,27.42~40.32,雄虫36.56~46.53),基部直径为1.61~2.78 μm,基窝宽度为3.23~5.05 μm。主要分布于触角鞭节各亚节的腹面、侧面和各亚节的连接处,鞭节背面数量很少,通常为1~2根,被鳞形感器包围(图2-C)。
毛形感器II,在椰子织蛾触角上分布数量少于毛形感器I,感器与触角表面的夹角为20°~45°,感器顶端稍稍弯曲,感器壁上有纵向纹路,基部也有明显的感器窝,端部钝圆。感受器的平均长度为14.4~26.17 μm,基部直径为1.4~2.2 μm,感器窝宽度为3.6~4.5 μm。主要分布于触角鞭节各亚节的侧面,腹面也有少量分布,通常与毛形感器I混生在一起(图2-D)。
(3)鳞形感器(Sensilla squamiformia)
鳞形感器一般存在于长鳞片的覆盖下,感器呈现细长柳叶状,尖端急尖,着生在感器窝内,感器表面有深的竖形条纹,感器的表面未发现有小孔。长度为29.14~32.8 μm,中部宽度为2.40~2.75 μm,基部窝宽度为1.7~2.2 μm,主要分布在鞭节各亚节背面节与节的链接处,或者鞭节中部。(见图2-E)
(4)锥形感器(Sensilla basiconca)
锥形感器呈现锥状,感器窝明显或无,有些有基座,表面有较深的纵纹、刻点和小孔,顶端急尖或钝尖。在椰子织蛾触角上分布有3种类型的感器,主要分布于触角的腹侧面。
事业单位的本质为,国有资产出资创立、主要运营方向带有浓郁的社会公益色彩,并受到党的直接领导与管控,事业单位主要为学校等教育事业单位、医院等卫生事业单位组成。而因事业单位的特殊性决定,党对其掌控力度的强弱至关重要,而政工工作作为党控制事业单位的主要工作内容与表现形式,极大增强了事业单位中党的实际控制能力。
锥形感器 I,感器表面有较深的纵纹和刻点,端部急尖,感器窝不明显;长度为11.6~18.2 μm,基部直径为1.2~2.8 μm;感器通常指向触角端部,分布于触角鞭节各亚节的侧腹面(图2-F)。
锥形感器II,感器表面有浅的“回”字纵纹、刻点和小孔,端部钝尖,有较深的感器窝;长度为24.1~26.7 μm,基部直径为1.7~2.5 μm;感器通常指向触角端部,触角鞭节各亚节腹面分布较多,侧面也有分布(图2-G)。
锥形感器Ⅲ,感器表面有浅的纵纹,接近基座有刻点;基部有基座,端部钝圆,感器窝不明显;长度为11.6~15.7 μm,基部直径为1.8~2.2 μm;感器与触角表面呈30°~45°夹角。主要分布于触角鞭节各亚节腹面侧面(见图2-H)。
(5)栓锥形感器(Sensilla styloconica)
栓锥形感器形似大拇指,有短而粗的基座,基座呈现圆柱形,顶端有凸出的芽锥,芽锥顶端有小孔,基座表面有纵向深纹汇集于基部。基座长度为12.7~15.6 μm,基座直径5.6~11.1 μm,芽锥高度1.44 μm,芽锥直径0.78 μm。该主要分布于鞭节各亚节的侧面或腹面的端部(图2-I)。
(6)腔锥感器(Sensilla coeloclnica)
腔锥感器由表皮下陷形成,中心具有锥形感器,感器的端部钝尖。按照感器的形状、大小和缘毛长短,可以分为两种:腔锥感器I(长缘毛);腔锥感器II(短缘毛)。雄虫未见到腔锥感器II。
腔锥感器II,感器周围被短缘毛聚拢,短缘毛呈短刺状。中心的锥形感器壁上比较小,顶端或壁具有小孔。中心的锥形感器长度为1.1~3.4 μm,直径为0.9~1.4 μm。主要分布于雌成虫触角鞭节各亚节的中部,通常为1个(图2-K)。
(7)Böhm氏鬃毛(Böhm bristles)
Böhm氏鬃毛是分布于柄节和梗节的基部感器。该感器直立于触角表面,与刺形感器相比短而尖,感器壁光滑无孔。长度为6.5~11.8 μm,基部直径0.8~1.3 μm,在柄节与梗节基部成簇着生,鞭节整个亚节均未观察到(图2-L)。
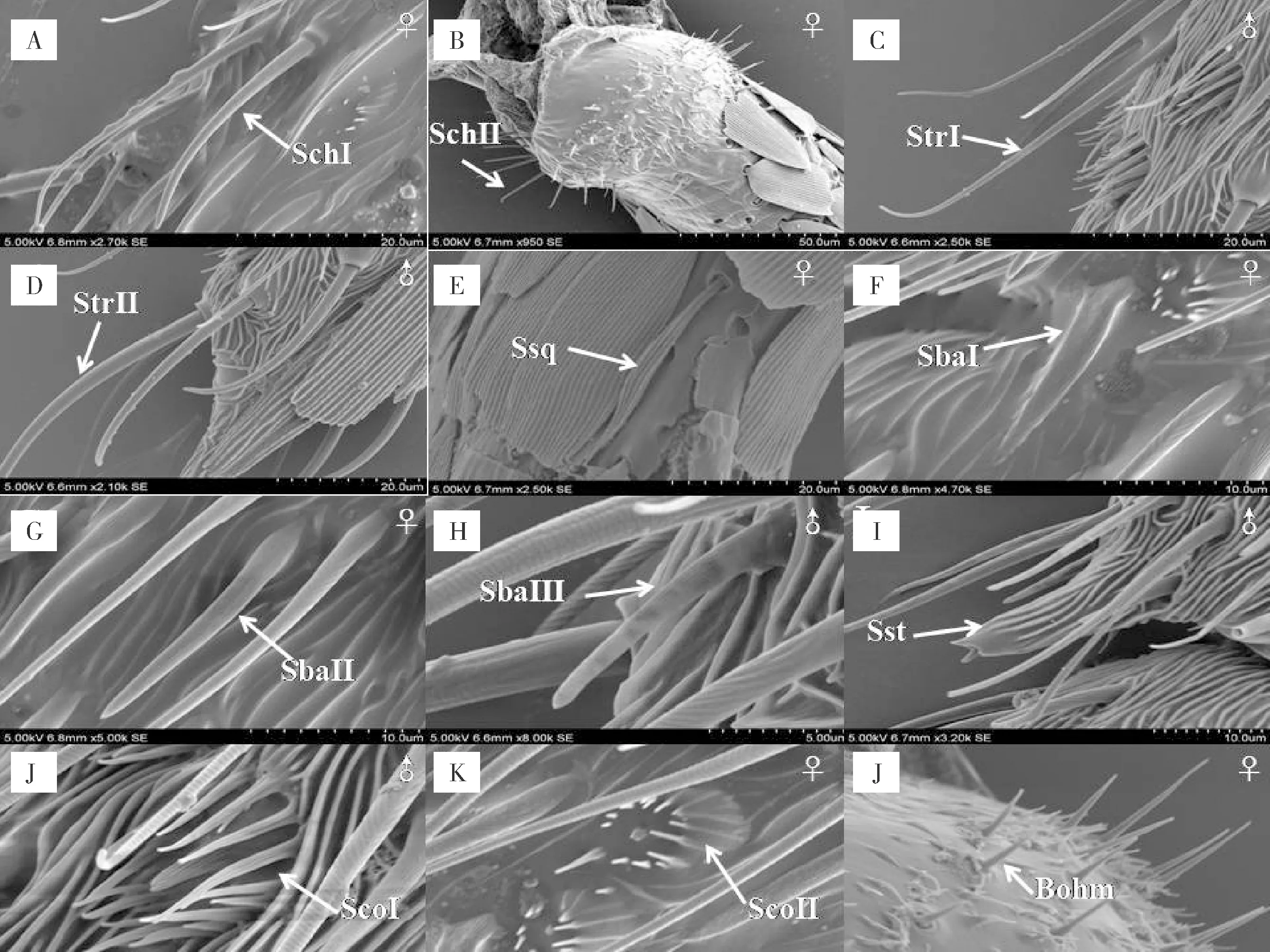
图2 椰子织蛾触角感器扫描电镜图Fig.2 Scanning electron micrograph of the antennal sensilla of Opisina arenosella注:A,SchI,刺形感器I;B,SchII,刺形感器II;C,StrI,毛形感器I;D,StrII,毛形感器II;E,Ssq,鳞形感器;F,SbaI,锥形感器I;G,SbaII,锥形感器II;H,SbaⅢ,锥形感器Ⅲ;I,Sst,栓锥形感器;J,ScoI,腔锥感器I;K,ScoII,腔锥感器II;L,Böhm,Böhm氏鬃毛。Note: A, SchI, sensilla chaetica I; B, SchII, sensilla chaetica II; C, StrI, sensilla trichodea I; D, StrII, sensilla trichodea II; E, Ssq, sensilla squamiformia; F, SbaI, sensilla styloconica I; G, SbaII, sensilla basiconca II; H, SbaⅢ, sensilla basiconca Ⅲ; I, Sst, sensilla styloconica; J, ScoI, sensilla coeloclnica I; K, ScoII, sensilla coeloclnica II; L, Böhm, Böhm bristles.
3 结论与讨论
触角是昆虫感觉的最重要的器官,具有嗅觉、味觉、机械和感受外界环境温湿度等功能,在昆虫寻找寄主和配偶,判断环境适合度过程中有着重要的作用(那杰等, 2008)。本研究发现椰子织蛾成虫触角上存在7种感器类型,共13个亚型。
椰子织蛾成虫触角上着生的7种感器,在雌虫和雄虫上都有发现,只有腔锥感器II在雄成虫中没有发现。Shanbhag等认为腔锥感器II的环形围栏状微毛具有保护内部着生感觉锥免受外界物理损伤的作用(Shanbhagetal., 1995)。另外,腔锥感器II作为嗅觉感受器官,通常被认为是在雌性寄主产卵选择上起作用(Hansson, 1995; Hunger and Steinbrecht, 1998)。本研究推测腔锥感器II在椰子织蛾雌雄成虫触角上的形态差异,可能与雌虫较雄虫因对寄主产卵选择的需求而更容易感受寄主挥发物的刺激有关。
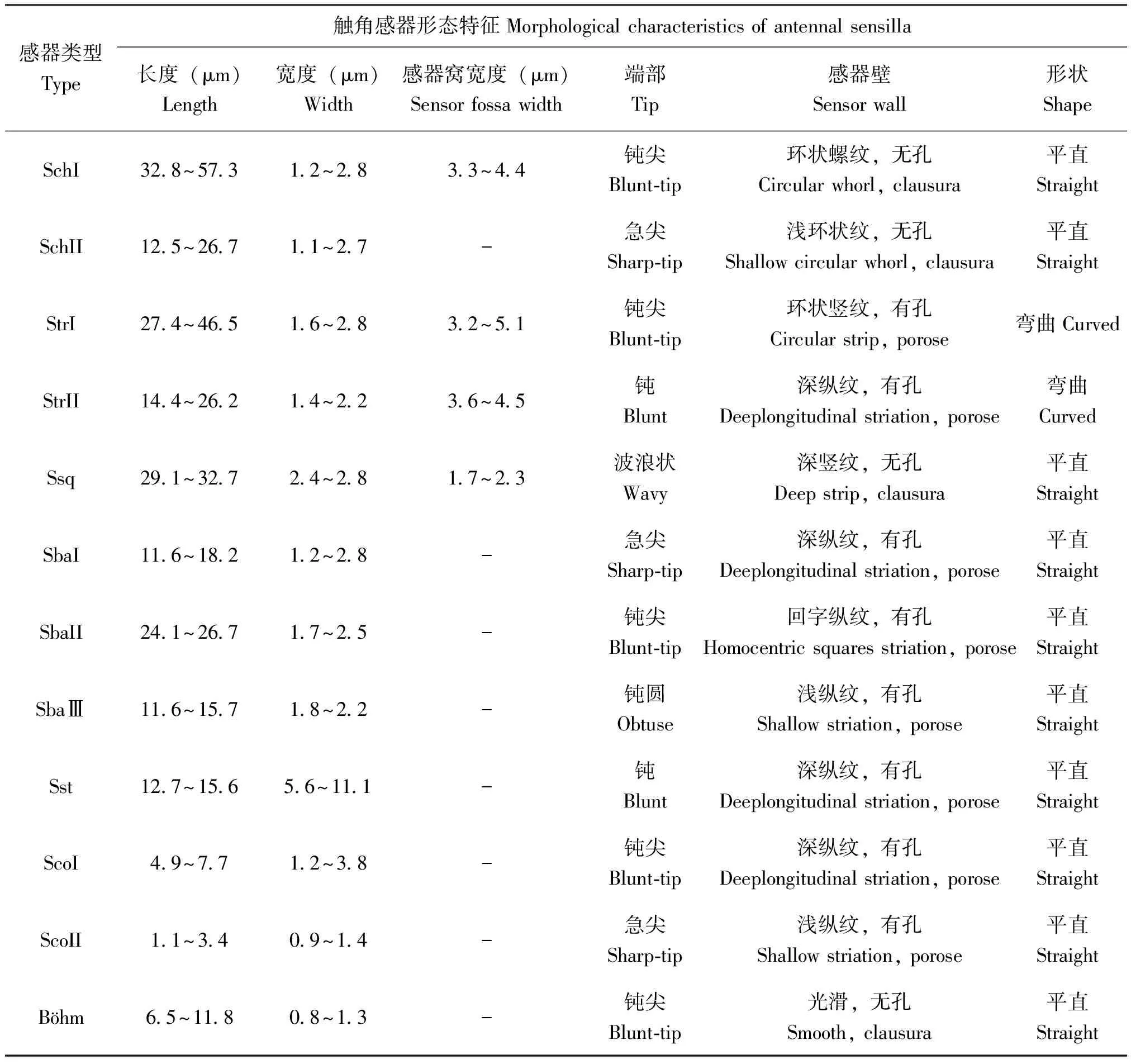
表3 椰子织蛾成虫触角上感器的类型
和鳞翅目其它昆虫的触角感器类型相比,刺形感器II为本研究在鳞翅目昆虫中新发现的一种感器,从长度、直径、宽度等指标来看,别的昆虫中均没有。刺形感器被认为是机械感器,用于保护触角、接触物品,感觉机械刺激的功能。椰子织蛾的刺形感器II与其他昆虫的刺形感器有区别,可能与椰子织蛾喜欢在棕榈植物的老叶上产卵的选择习性有关。另外,没有在椰子织蛾成虫触角上发现卷蛾科梨小食心虫Grapholithamolesta触角上的耳形感器、荔枝异性卷蛾Cryptophlebiaombrodelta触角上的板形感器和黄斑长翅卷蛾Aclerisfimbriana触角上的钟形感器(韩桂彪和马瑞燕, 1996)。耳形感器在夜蛾科和卷蛾科的部分种类研究中被发现,据推断,这类感器与成虫探测植物气味有关,有报道在卷蛾科的C.pomonella上发现此感器起到感受微量信息素的功能(Denetal., 1978; Andersonetal., 1995; Zebitz, 1998; Rohetal., 2016)。板形感器被认为其功能为感受红外辐射,钟形感器对气味、二氧化碳、温湿度敏感。通常,不同习性的昆虫在触角感器上有差异,如散居型和群居型。散居型的昆虫比群居型拥有更多的嗅觉感器。另外,生存环境、天敌数量等都与触角感器的类型和数量有关(那杰等, 2008)。
毛形感器通常感器壁上由小孔或者小沟同外界空气连接,具有嗅觉功能。在梨小食心虫、桃小食心虫、小地老虎、棉铃虫等鳞翅目昆虫中均有发现。锥形感器是比较有特色的一类感器,具有嗅觉和味觉功能,这和桃小食心虫(宫田睿等, 2016)、梨小食心虫(郭永福等, 2018)、甜菜夜蛾(徐进等, 2018)的研究结果一致。栓锥感器具有嗅觉和感受温、湿度的功能,腔锥感器在椰子织蛾上分布比其它鳞翅目昆虫多,具有嗅觉和感受温度的功能。鳞形感器在椰子织蛾触角上发现数量最少,向玉勇等(2016)报道该感器为机械感器,只有支撑功能,也有学者报道其具有嗅觉功能。Böhm氏鬃毛只分布于触角的柄节与梗节的基部,在鞭节并未发现,这一结果与鳞翅目很多昆虫触角感器的分布一致(Liuetal., 2018)。有研究者还认为其具有感受温度和湿度等物理刺激的功能。关于Böhm氏鬃毛是否真的具有这一功能还需要继续研究。
椰子织蛾触角表面密被鳞片,会与鳞形感器混淆,也有学者直接将其命名为鳞形感器,本研究认为报道有误,主要原因是经过氢氧化钠处理,鳞片自然脱落,因此其不具有任何感器的功能。关于感器的分类,目前国内外并无统一标准,主要原因是昆虫种类多,感器类型相差过大,并且在未来可能会有新的发现。在今后的研究中,应当使用新的科学技术例如免疫组化技术,投射电镜技术、转录组测序技术等,对触角感器功能重新进行研究,如果能确定感器真正的功能,这将对害虫的防治有很大意义。
