访花昆虫野蚜蝇线粒体基因组结构分析
2021-08-13程梦迪曹春桥
闫 艳,程梦迪,曹春桥,李 虎
(陕西理工大学生物科学与工程学院,陕西汉中 723000)
野蚜蝇Syrphustorvus(Osten-Sacken, 1875)是双翅目Diptera蚜蝇科Syrphidae常见昆虫,其成虫和幼虫在农林业生产过程中都扮演重要角色。成虫主要外形特征为头和中胸背板暗黑色;小盾片黄色;腹部黑色,第2背板具1对黄斑,第3、4背板各具黄色横带,第4、5背板后缘黄色(黄春梅等,2013),具有访花习性,常见于林间野花、芸薹属植物及禾本科植物附近(Fazlullahetal., 2018),是重要的传粉昆虫;幼虫为蛆,可捕食多种植物上的蚜虫,在1960年Brown和Clark的研究中,野蚜蝇幼虫是冷杉球蚜Adelgespiceae的主要天敌之一。该种在古北区、新北区、东洋区、新热带区和澳洲区均有分布(霍科科等,2006)。
蚜蝇的形态和体色多变,生物学特征复杂,不同学者提出了不同的分类系统,现行的3亚科分类系统:蚜蝇亚科Syrphinae、管蚜蝇亚科Eristalinae和巢穴蚜蝇亚科Microdontinae,被大多数学者接受。近些年来有些学者利用线粒体与细胞核的基因片段构建系统发育树,讨论蚜蝇科分类系统,把蚜蝇科中缩颜蚜蝇提为亚科水平,将蚜蝇科划分为4亚科(蚜蝇亚科、管蚜蝇亚科、巢穴蚜蝇亚科和缩颜蚜蝇亚科Pipizinae)(Mengualetal., 2015)。线粒体基因组由于其母系遗传、分子较小、突变率高等特点,被广泛用于昆虫的鉴定、种群遗传、物种进化、系统发育树构建的研究中(Cameron, 2014)。
本研究对野蚜蝇的线粒体基因组全序列进行测序组装、基因注释,分析其基因组的基本结构特征,进一步丰富了蚜蝇科线粒体基因组学研究;利用野蚜蝇线粒体全序列并联合已公布的21种蚜蝇科线粒体全序列,重新构建蚜蝇科系统发育树,尝试对蚜蝇科内部分类群间的系统发育关系进行讨论分析。
1 材料与方法
1.1 标本采集与保存
本实验所用的野蚜蝇于2019年3月23日采自陕西省汉中市南郑区大汉山(106.93° E, 33.97° N),无水乙醇浸泡并保存于-20℃的冰箱。
1.2 DNA提取与基因组测序
取野蚜蝇胸部和腿部肌肉组织,用无菌研磨棒将其研磨成粉末状后严格按照基因组DNA提取试剂盒中使用手册(天根生化科技有限公司)进行基因组DNA提取,检验合格(使用超微量紫外分光光度计检测DNA浓度>25 mol/μL)后将基因组送至公司于Illumina NovaSeq 6000测序平台进行高通量测序,测序模式为150 PE,每样品产生6 GB数据,每样本数据量不低于目标量的95%,平均Q≥85%。
1.3 PCR扩增与测序
COXI基因的片段序列(约600 bp)通过构建50 μL体系进行PCR扩增,其中基因组DNA 5 μL,TaqDNA聚合酶25 μL,上下游通用引物LCO1490(GGTCAACAAATAATAAAGATATTGGG)和HCO2198(TAAACTTCAGGGTGACCAAAAAATCA)和Mg2+各1 μL,超纯水17 μL。
PCR扩增的反应条件:94℃预变性4 min一个循环,94℃变性30 s,45℃退火30 s,72℃延伸45 s,35个循环,最后在72℃下后延伸10 min。PCR产物使用琼脂糖凝胶电泳检测合格后,进行测序。
1.4 序列拼接、注释与分析
利用Geneious R9软件,使用COXI基因的片段序列作为诱饵进行序列组装,并对获得的线粒体基因组全序列进行注释,利用ARWEN(Laslett and Canbäck, 2008)预测22个tRNA位置和二级结构,利用ClustalX比对意大利蜜蜂Apismellifera、烟草天蛾Manducasexta和横带刺茎叶蝉Taharanafasciana的16SrRNAs和12SrRNAs预测野蚜蝇的2个 rRNAs二级结构(Gillespieetal., 2006;Cameron and Whiting, 2008;Wangetal., 2017)。
1.5 系统发育树的构建
本研究收集了NCBI数据库中已公布的21条蚜蝇科和2条缟蝇科物种线粒体全基因组序列(表1)。利用包括野蚜蝇在内的24个物种线粒体基因的13个PCGs序列,使用比对网站http://pc16141.mncn.csic.es/index_v4.html(Abascaletal., 2010)将13个PCGs序列分别比对后在MEGA7中串联(Kumaretal., 2016),在分区网站https://cushion3.sdsc.edu/portal2/login!input. action(Milleretal., 2010)上,使用PatitionFinder 2 on XSEDE工具选取适合PCG数据集的最佳分区,BI树在建树网站https://cushion3.sdsc.edu/portal2/login!input.action运行5 000代,收敛值为0;使用IQ-tree(Nguyenetal., 2015)构建ML树,参数为Bootstrap analysis: Ultrafast, Number of bootstrap alignments: 1 000, Maximum iterations: 1 000, Minimum correlation: 0.99,重复1 000次。

表1 本研究所用物种完整线粒体基因组信息
2 结果与分析
2.1 基因组结构
野蚜蝇线粒体基因组全长16 444 bp,总A+T含量为80.6%,各类型基因的AT含量均大于78%,表现出较强AT偏向性,详见表2。线粒体基因组包括13个PCGs,22个tRNAs,2个rRNAs和D-loop区。基因间隔共19处,最长的间隔位于tRNA-Tyr和COXI之间。基因重叠共有12处,最长基因重叠区位于ATP8和ATP6之间。37个基因中,4个tRNAs和9个PCGs位于J链,其余8个tRNAs、4个PCGs和2个rRNAs位于N链(表3)。

表2 野蚜蝇线粒体基因组碱基组成及偏向性
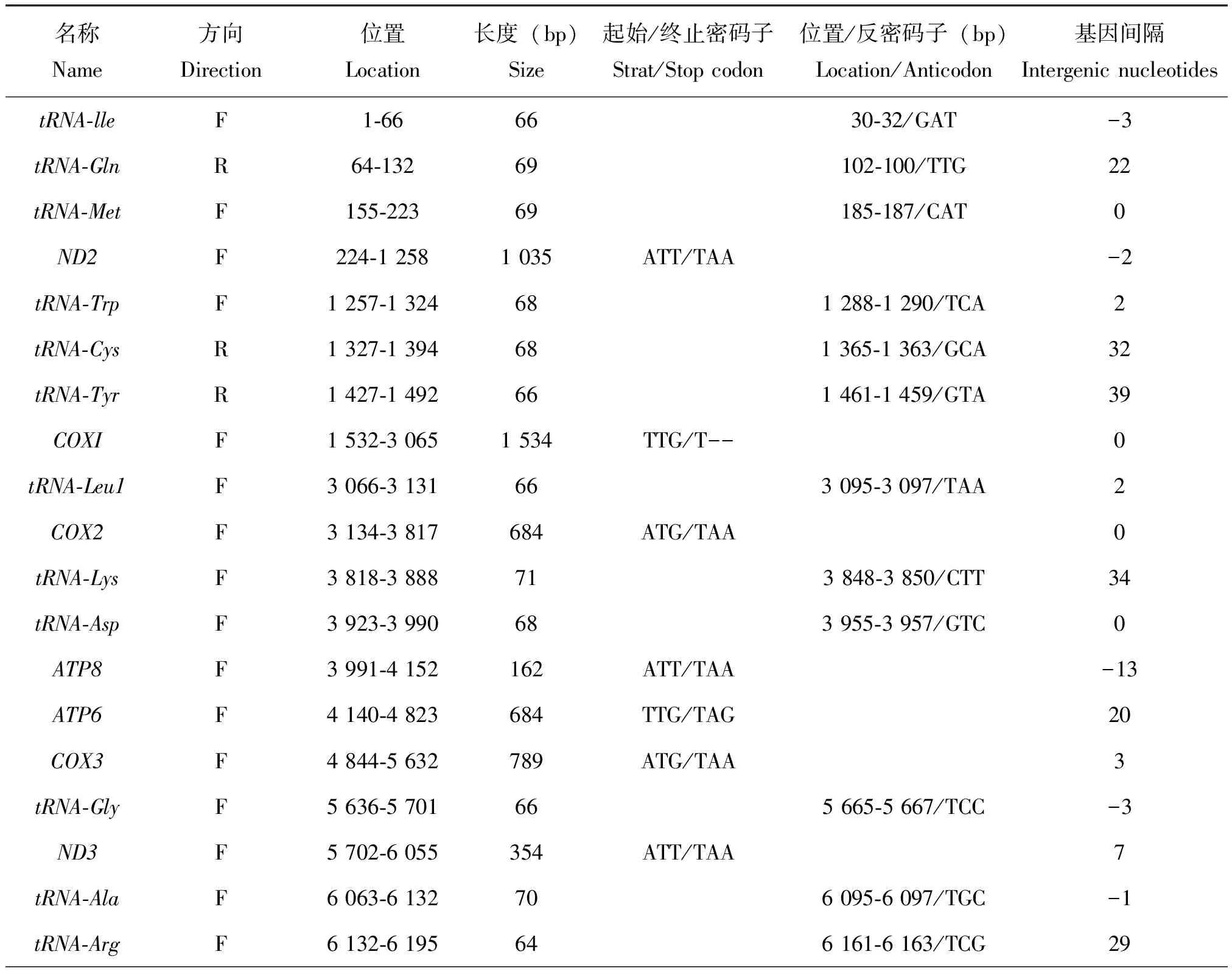
表3 野蚜蝇线粒体基因组组成

续表3 Continued table 3
2.2 蛋白编码基因
野蚜蝇PCGs序列长度为11 230 bp,A+T含量为78.4%,AT skew为-0.142,T的含量高于A的含量。以ATN为起始密码子的基因有9个,其中ND2、ND5和ND6以ATT为起始密码子,ND4L、COX2和ND3以ATA为起始密码子,COX3、ND4和Cytb以ATG为起始密码子;COXI、ATP8、ATP6、和ND1以TTG为起始密码子。COXI的终止密码子为不完整T--,ATP6的终止密码子为TAG,其余11个PCGs的终止密码子都为TAA(表3)。
野蚜蝇PCGs序列共编码5 480个氨基酸,其中使用频率最高的密码子是AAA共437个,而相对密码子使用频率最高的密码子为UUA,频率为2.92;Leu是使用频率最高的氨基酸。从密码子使用情况来看,使用频率较高的密码子,如AAA、AAU和AUU等,它们的碱基大部分都由A、T组成(图1),这可能是PCGs序列中AT偏斜大于GC偏斜的原因。

图1 蛋白编码基因相对同义密码子使用和密码子数量Fig.1 Relative synonymous codon usage(RSCU)and codon number of PCGs in Syrphus torvus mitogenome
2.3 tRNAs基因与rRNAs基因
22个tRNAs基因总长度为1 472 bp,单个基因长度从64 bp(tRNA-Arg)到72 bp(tRNA-Val)不等。22个tRNAs的二级结构都是典型的三叶草结构(图2),出现了37对错配,其中DHU臂上有18对,反密码子臂上有10对,氨基酸接受臂上有5对,TΨC上有4对,其中G-T错配数量最多,共有17对。
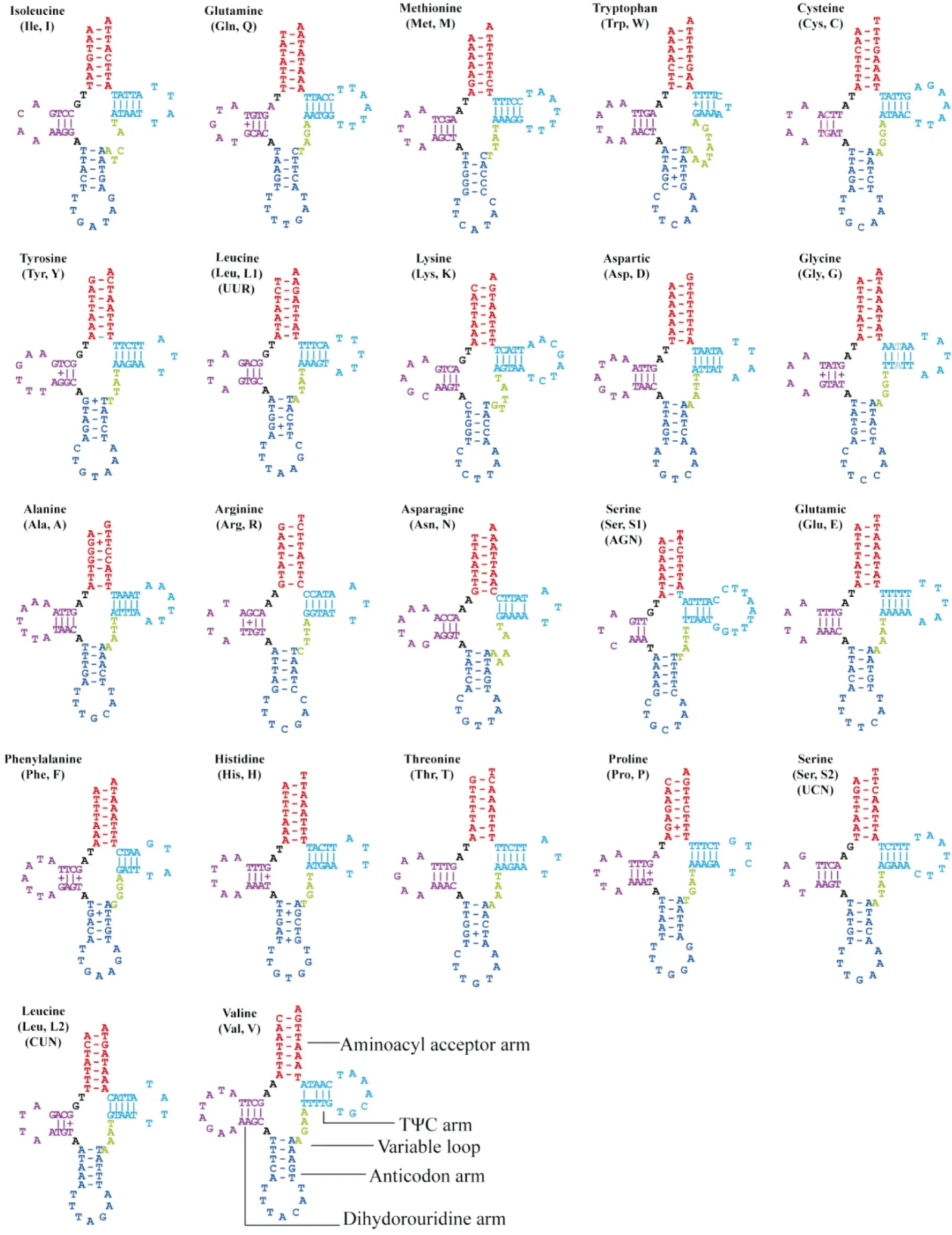
图2 野蚜蝇22个tRNAs的二级结构Fig.2 Secondary structure of the 22 tRNAs genes of Syrphus torvus
两个rRNAs都位于N链,16SrRNA位于tRNA-Leu2和tRNA-Val之间,长度为1 342 bp;12SrRNA位于tRNA-Val和D-loop之间,长度为791 bp。16SrRNA的二级结构由I、II、III、IV、V和VI 6个区域组成(节肢动物第Ⅲ区域缺失),44个螺旋结构(图3);12SrRNA二级结构由I、II和III 3个区域组成,24个螺旋结构(图4)。

图3 野蚜蝇16S rRNA二级结构预测Fig.3 Predicted secondary structure of the 16S rRNA of the mitogenome of Syrphus torvus注:破折号(-)代表Watson-Crick配对,点(·)代表GU配对。Note:Dashes(-)indicated Watson-Crick base pairing, hereas GU pairs were connected by dots(·).
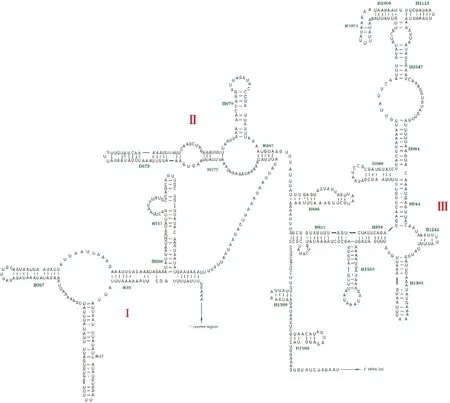
图4 野蚜蝇12S rRNA二级结构预测Fig.4 Predicted secondary structure of the 12S rRNA of the mitogenome of Syrphus torvus
2.4 非编码区与重叠区
野蚜蝇线粒体基因组除D-loop区外,有19个非编码区,长度在2~39 bp之间;9个重叠区,长度在1~13之间。
D-loop区位于12SrRNA与tRNA-Ile之间,长度为1 379 bp。控制区的A+T含量在所有基因中含量最高,为92.2%。在该区找到了4个长度大于15 bp的重复序列和3个茎环结构,详见图5。

图5 野蚜蝇D-loop的重复序列和茎环结构Fig.5 Repetitive sequences and the stem-loop structures in the D-loop of Syrphus torvus注:蓝色表示重复,黄色表示茎环结构。Note:Bule for repetitions and yellow for stem-loops.
2.5 系统发育关系
缟蝇科2个物种作为外群,22种蚜蝇(含本研究测序的野蚜蝇)为内群。选择线粒体全序列的13个PCGs序列构建数据集,使用BI和ML两个方法构建系统发育树(图6~图7)。在两个系统发育树中,野蚜蝇聚在了蚜蝇亚科Syrphinae,始终与黑足蚜蝇Syrphusvitripennis聚成一支,互为姊妹种;蚜蝇亚科物种均聚为一支,且支持度均很高,表现出单系性,管蚜蝇亚科Eristalinae物种没有聚成一支。
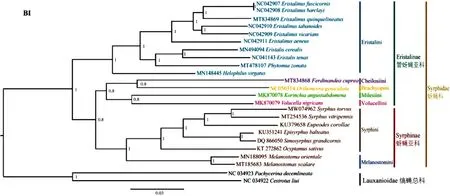
图6 基于13个蛋白编码基因构建的贝叶斯树(BI)Fig.6 BI tree built based on 13 PCGs
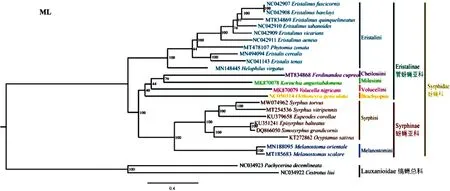
图7 基于13个蛋白编码基因构建的最大似然树Fig.7 ML tree built based on 13 PCGs
在蚜蝇亚科中,墨蚜蝇族Melanostomini(东方墨蚜蝇Melanostomaorientale+梯斑墨蚜蝇Melanostomascalare)聚为姊妹种并与蚜蝇族Syrphini互为姊妹群关系;蚜蝇族中蚜蝇属(野蚜蝇Syrphustorvu+黑足蚜蝇Syrphusvitripennis)聚为一支;蚜蝇亚科中(((黑带蚜蝇Episyrphusbalteatus+大角类蚜蝇Simosyrphusgrandicornis)+Ocyptamussativus)+大灰优蚜蝇Eupeodescorollae),它们分属蚜蝇族不同属,在系统发育树中的聚类与前人研究结果一致(Puetal., 2017; Li, 2019; Li and Li, 2019、2020)。管蚜蝇亚科中管蚜蝇族Eristalini单独聚为一支,而黑蚜蝇族Cheilosiini、迷蚜蝇族Milesiini、蜂蚜蝇族Volucellini和Brachyopini聚在一起并与蚜蝇亚科聚为姊妹群关系。管蚜蝇族中斑眼蚜蝇属Eristalinus物种聚为一支,管蚜蝇属Eristalis两个物种聚为姊妹关系,与Yan等(2020)和Li等(2020)的研究结果一致。
3 结论与讨论
本研究获得了野蚜蝇Syrphustorvus线粒体基因组全序列,长度为16 444 bp,A+T含量为80.6%,表现出明显的AT偏向性;野蚜蝇线粒体包含22个具典型三叶草结构的tRNAs基因,长度在64~72 bp之间;目前蚜蝇科物种线粒体rRNAs基因的二级结构还没有被预测,由于双翅目Diptera昆虫中的rRNA基因二级结构都没有被预测,本研究参照膜翅目Hymenoptera、鳞翅目Lepidoptera和同翅目Homoptera已预测的16SrRNA和12SrRNA二级结构,使用ClustalX预测野蚜蝇rRNAs二级结构。
野蚜蝇D-loop区的A+T含量最高(92.2%),同时也发现存在一些重复序列和茎环结构,这些结构可能在控制DNA甲基化、基因转录和复制方面发挥作用(Huetal., 2019)。在本研究所涉及的22个蚜蝇的基因间隔区与重叠区中发现了6个保守区域:区域1:位于tRNA-Trp与tRNA-Cys之间(AAGTCTTA);区域2:位于tRNA-Tyr与COXI之间(TCAACTAATCATAAAGAT);区域3:位于RNA-Lys和tRNA-Asp之间(Poly A);区域4:位于ATP8与ATP6之间(ATGATAA);区域5:位于ND4和ND4L之间(TTAACAT);区域6:位于ND1与tRNA-Leu2之间(ATAAAACAAC)。区域1、3、4和6在蚜蝇亚科物种中被发现;区域2、4和5在管蚜蝇亚科物种中被发现。此外,区域4不仅发现存在于蚜蝇科,在半翅目Hemiptera一些物种中也有发现,该区被认为是在翻译时作为一个双顺反子使用(Xuetal., 2019)。这些保守区域在控制PCGs序列长度和转录等方面有重要作用(Cameron and Whiting, 2008)。
BI和ML系统发育树显示,蚜蝇亚科Syrphinae的分支都很稳定,其单系性也被大多数学者认可(Puetal., 2017; Paulietal., 2018;Li, 2019)。而管蚜蝇亚科Eristalinae的系统发育关系还存在一定的争议,Ståhls等(2003)利用形态学与分子相结合的方法构建系统发育树证实管蚜蝇亚科为单系;然而,近年来,Skevington和Yeates(2000)、Mengual等(2015)、Young等(2016)和Pauli等人(2018)的研究结果显示管蚜蝇亚科为并系,本研究结果也支持这一结论。
管蚜蝇亚科内部的系统发育关系存在一定争议,如斑眼蚜蝇属Eristalinus,在BI树中,其拓扑结构(图6)如下:(((E.fuscicornis+E.barclayi)+(E.tabanoides+E.vicarians))+黑色斑眼蚜蝇Eristalinusaeneus),与Sonet等(2019)和Yan等(2020)的研究结果一致;而在ML拓扑结构(图7)中:(((E.fuscicornis+E.barclayi)+E.tabanoides)+E.vicarians)+黑色斑眼蚜蝇Eristalinusaeneus),这与Li等(2019, 2020)研究结果相同;黑蚜蝇族Cheilosiini、迷蚜蝇族Milesiini、蜂蚜蝇族Volucellini和Brachyopini各只有一个代表种,系统发育关系不稳定,在BI拓扑结构中,黑蜂蚜蝇Volucellanigricans与狭腹斜环蚜蝇Korinchiaangustiabdomena聚为姊妹种,符合Li(2019)的研究结果,而在ML拓扑结构中,黑蜂蚜蝇和狭腹斜环蚜蝇没有聚为一支,与Li等(2020)的研究结果一致。
本研究得到的两个蚜蝇科拓扑结构,蚜蝇亚科Syrphinae始终聚成一支,其单系性得到了验证;而管蚜蝇亚科Eristalinae并未聚成一支,可能为并系,支持前人的研究结果,但这一结论仍需进一步验证,后期应加大基因组测序力度,补充大量蚜蝇科物种基因组后,进一步深刻理解、阐明蚜蝇科系统发育关系。
