氮钙协同对杨树生长、光合特性及叶绿素荧光的影响
2021-08-11翁小航周永斌任城帅张淞著朱文旭刘丽颖杨成超
翁小航,李 慧,周永斌,任城帅,张淞著,朱文旭,刘丽颖 ,杨成超
(1.沈阳农业大学林学院,沈阳110161; 2.辽河平原生态站,辽宁昌图112500; 3.辽宁省杨树研究所,辽宁盖州115200)
杨树不仅是我国重要的经济林速生树种,也是我国干旱半干旱地区的主要生态防护树种之一[1]。本试验中的“中辽一号”杨(Populus×canadensis cv.‘zhongliao 1’)是在20 世纪70 年代末,由黄东森等[2]以I-69 杨、欧黑(Populus.nigra)、箭黑(Popolus.thevestina×Populus.nigra)、青阳(Populus.cathayana)等为亲本杂交而成的无性系品种,具有干型好、生长快及抗病虫能力强等优势,所以被广泛应用在辽宁地区的护路林、固沙林、农田防护林及农牧防护林中。由于降雨缺乏导致水土流失、养分不均衡问题日益严重,“中辽一号”杨长势也出现了衰退现象,不仅造成杨树人工林或混交林在某地区的生态功能降低,还造成了严重的经济损失[3]。
氮、钙作为植物生长发育必需的营养矿质元素,在植物生长发育过程中起着至关重要的作用[4]。氮素是植物生长发育必需的营养元素,由于氮可以合成捕获光合电子传递的相关蛋白及酶[5],所以氮素含量的高低直接影响了光合作用的强弱。施加适量的氮素可以促进植物叶片中叶绿素的累积,使植物体净光合速率增强,有助于有机物的积累,提高植物干重[6-8];而氮含量过高,则会对植物细胞产生胁迫作用,抑制植物生长[9]。钙不仅是植物生长的必需营养元素,还能对植物的各项生理活动起到调节作用,参与植物的光合作用,对植物的抗逆性产生显著影响。当外界供给的Ca2+低于植物体所需的正常范围时,植物的生长及光合会受到影响,减弱植物的抗逆性,引起各种植物病害,如白菜烧心病[10-11]等;而Ca2+过多则会产生毒害作用,导致气孔关闭,叶绿体内膜受损,破坏类囊体薄膜,干扰光合作用,抑制植株生长[12]。
目前,关于氮钙协同对植物生长发育的影响已在黄瓜[13]、、青花菜[14]、番茄[15]、小麦[16]和设施油桃[17]等农产品、经济作物上取得初步研究进展,但关于杨树之类的速生乔木树种的研究还尚未有进展。植物想要达到较高的生物量,其光合作用必须维持在较高的水平,叶绿素荧光参数可以反映植物叶片光合作用的强弱,从而推断植物的生长状况。自然界土壤中钙元素具有较高的空间异质性,并且氮沉降问题日益严峻,严重影响到植物的分布及生长。因此,本试验以杨树为研究对象,进行不同氮钙协同处理,从生物量、光合参数和叶绿素荧光方面进行研究,为进一步研究氮钙协同对杨树生长发育特性的影响和杨树人工林的培育提供理论依据。
1 材料与方法
1.1 杨树幼苗的培育
试验于2018 年4~8 月在沈阳农业大学后山实验基地进行。选用1 年生、株高与地径大小一致的杨树扦插苗进行水培,共10 个处理,每个处理3 个重复。2018 年4 月24 日开始对其进行水培,使其适应一段时间,待幼苗生根后(5月16日)对其进行氮处理和钙处理,5~7月每7d更换1次营养液,直至7月9日收苗。
1.2 外源钙及外源氮的处理
营养液根据谢晓梅[18]砂培营养液配方,用蒸馏水配置,用NaOH 调节pH 值为5~6。试验设定5个Ca2+浓度水平:0,2.5,5,10,20mmol·L-1;2 个N 水平:5 mmol·L-1和20mmol·L-1。钙梯度由无水CaCl2提供,氮梯度由Na-NO3和NH4Cl 按照1∶1 的浓度进行配制。其他大量元素分别由KCl、MgSO4·7H2O、KH2PO4、EDTA-Na2和FeSO4·7H2O组成,保证各处理中的P、K、Mg和S的含量分别为1.0,6.0,2.0,2.0mmol·L-1。
1.3 测量方法及指标
1.3.1 生物量的测定 水培结束后将幼苗取出,并保证植株的完整性,用蒸馏水将整株幼苗清洗干净后,将杨树幼苗分为根、茎、叶3部分,用剪枝剪剪断,并用信封装好,做好标记,在烘箱中先用105℃杀青0.5h后,再将烘箱调节至80℃,烘干至恒重,使用千分之一天平称量各部分干重,并保留两位小数。
1.3.2 光合参数的测定 光合参数使用Li-6400 光合仪测定:在杨树幼苗生长高峰期(2018 年6 月),在晴朗天气条件下的每日9∶00~11∶00,使用便携式光合仪Li-6400(LI-COR,Inc,USA)测定杨树幼苗叶片的净光合速率(Pn),气孔导度(GS)和蒸腾速率(Tr)等光合参数指标,将有效光辐射强度(PAR)控制在1000μmol·m-2·s-1,每个处理选取3株幼苗,每株重复3次取平均值。
1.3.3 叶绿素荧光参数的测定 采用便携式脉冲调制荧光仪(OPTI-SCIENCES, OS5p+),于9∶00~11∶00, 选择从上往下数经过充分光照的、完全展开的第2 片叶子,先用暗适应夹夹住叶片进行20min 的暗适应,然后开始测定不同氮钙协同处理下杨树叶片的最大光化学效率(Fv/Fm)和潜在光化学效率(Fv/F0)。
1.3.4 数据处理 利用Excel2017和SPSS22.0软件对试验数据统计分析。图表中数据为3次重复的平均值±标准误所得,并采用单因素方差分析法和Duncan(新复极差法)法对数据进行差异性分析和显著性比较,误差线用标准误差表示,图表中不同字母表示不同氮钙协同处理间各指标差异达0.05显著水平。
2 结果与分析
2.1 氮钙协同对杨树幼苗根、茎、叶和总的生物量的影响
由表1可知,当氮处理相同时,杨树幼苗的根、茎、叶和总的生物量均随着钙浓度的升高呈现先升高后降低的趋势。在低氮处理下(5mmol·L-1),当Ca2+浓度为5mmol·L-1时,根、茎、叶和总生物量达到最大值,分别为1.61,1.69,0.89,4.20g,与不施钙时相比存在显著差异(p<0.05)。在高氮的处理下(20mmol·L-1),当Ca2+浓度为10mmol·L-1时,根、茎、叶和总的生物量达值最大,分别为1.45,1.24,0.86,3.56g,与不施钙时相比差异显著(p<0.05)。当钙处理相同时,比较氮对总生物量指标的影响发现,当Ca2+浓度在0~10mmol·L-1时,低氮处理下(5mmol·L-1)的总生物量大于高氮条件下(20mmol·L-1)的总生物量;当Ca2+浓度为10mmol·L-1时,不同氮处理下的总生物量差异不显著;而当Ca2+浓度为20mmol·L-1时,低氮处理下(5mmol·L-1)的总生物量小于高氮条件下(20mmol·L-1)的总生物量。因此氮钙协同显著影响杨树幼苗的生物量,除低氮(5mmol·L-1)、高钙(20mmol·L-1)对杨树幼苗生长产生严重胁迫外,其余处理均提高了杨树幼苗的根、茎、叶和总生物量,并且在5mmol·L-1的氮水平下添加5mmol·L-1的钙水平时,杨树幼苗的生长达到最优,并与其他处理差异显著(p<0.05)。
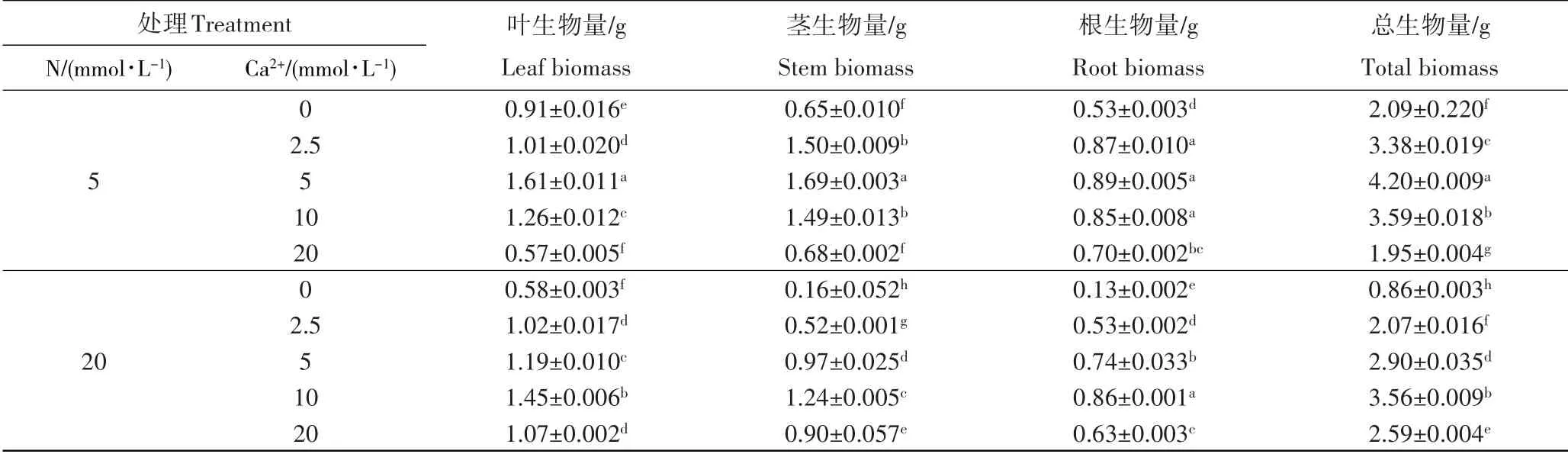
表1 氮钙协同对杨树幼苗根、茎、叶和总生物量的影响Table 1 The biomass of root, stem, leaf and total biomass of poplar seedlings under different nitrogen-calcium treatment
2.2 氮钙协同对杨树幼苗光合特征的影响
由图1 可知,当氮处理相同时,杨树幼苗的净光合速率、气孔导度和蒸腾速率均随着钙浓度的升高呈现先升高后降低的趋势。在低氮处理下(5mmol·L-1),当钙浓度为5mmol·L-1时,净光合速率、蒸腾速率和气孔导度均达到最大值,分别为5.96μmol·m-2·s-1、0.093mol·m-2·s-1和5.38mmol·m-2,与不施钙时相比存在显著差异(p<0.05)。在高氮处理下(20mmol·L-1),当钙浓度为10mmol·L-1时,杨树叶片的净光合速率、气孔导度、蒸腾速率达到最大值,分别为5.17μmol·m-2·s-1、0.088mol·m-2·s-1和4.23mmol·m-2,与不施钙时相比差异显著(p<0.05)。当钙处理相同时,比较氮处理对净光合速率、气孔导度和蒸腾速率的影响发现,当钙浓度小于10mmol·L-1时,高氮处理下(20mmol·L-1)的杨树叶片的净光合速率、气孔导度和蒸腾速率均低于低氮处理(5mmol·L-1);当钙浓度为10mmol·L-1时,高氮处理下(20mmol·L-1)的杨树幼苗叶片的净光合速率、气孔导度和蒸腾速率均显著高于低氮处理(20mmol·L-1);当钙浓度为20mmol·L-1时,高氮处理(20mmol·L-1)与低氮处理下(5mmol·L-1)的净光合速率、气孔导度与蒸腾速率均无显著差异。因此氮钙协同对杨树幼苗的光合作用影响显著,除高钙(20mmol·L-1)对杨树幼苗生长产生抑制外,其余处理均显著提高了杨树幼苗的光合参数,并且在5mmol·L-1的氮水平下添加5mmol·L-1的钙水平时,杨树幼苗的净光合速率、蒸腾速率和气孔导度达到最优,且与其他处理之间的差异均达到显著水平(p<0.05)。

图1 氮钙协同对杨树幼苗叶片光合作用的影响Figure 1 Effects of nitrogen-calcium treatment on photosynthesis of poplar seedling leaves
2.3 氮钙协同对杨树幼苗叶片光合系统Ⅱ的影响
由图2可知,当氮处理相同时,随着钙浓度的升高杨树幼苗PSⅡ的潜在光化学效率(Fv/F0)和最大光化学效率(Fv/Fm)均呈现出先升高后降低的趋势。在低氮处理下(5mmol·L-1),当钙浓度为5mmol·L-1时,其Fv/F0和Fv/Fm均达到最大值,与钙处理为2.5mmol·L-1和10mmol·L-1时的Fv/F0和Fv/Fm不存在显著差异,与钙处理为0 和20mmol·L-1时的Fv/F0和Fv/Fm存在显著差异(p<0.05)。高氮处理下(20mmol·L-1),当钙浓度为10mmol·L-1时,Fv/F0和Fv/Fm达到最大值,并与其他钙处理呈现显著性差异(p<0.05)。当钙处理相同时,比较氮对杨树的潜在光化学效率和最大光化学效率的影响发现,杨树低氮处理下低PSⅡ的Fv/F0和Fv/Fm均大于高氮处理,且存在显著差异(p<0.05)。氮钙协同条件下,当氮浓度为5mmol·L-1,钙浓度为5mmol·L-1时,PSⅡ的Fv/F0和Fv/Fm达到最大,当氮浓度为20mmol·L-1时添加10mmol·L-1的钙处理时达到最大值,与其他处理差异显著(p<0.05)。除此之外的其他处理下的杨树的最大光化学效率Fv/Fm均小于0.8,说明这些处理均对杨树幼苗的生长产生胁迫。

图2 氮钙协同对杨树幼苗潜在光化学效率和最大光化学效率的影响Figure 2 Effectof nitrogen-calcium treatment on potential photochemical efficiency and maximum photochemical efficiency of poplar seedlings
3 讨论与结论
氮、钙是植物生长发育所必需的大量元素,对植物的生长及光合特性起着不可或缺的作用。适量的钙处理能够有效促进植物的生长发育,改善光合代谢,使植物叶片保持较高的光合性能[19],有效稳定细胞结构、平衡离子状态维持细胞功能提高抗逆性[20]。同时,钙还可以促进植物体内可溶性糖及淀粉的转化和利用,促进植物体对其他营养元素的吸收,使细胞代谢过程维持在正常水平[21],确保植物能过进行正常的生长及生理活动。氮素是组成植物蛋白质和核酸的重要组成部分,叶绿素、蛋白质和多种维生素也是由大量氮元素构成[22]。而且氮素也是限制陆地生态系统生产力的重要营养元素之一。有研究表明适当的氮肥可促进根生长;而含氮量过多则会造成根系减少,根重降低,影响植物的生长,如冬小麦[23];当氮浓度为0.2g·kg-1时,菠菜的生物量会随着钙浓度的升高而增加[24],该结论与本试验结果相似。本试验表明,同氮浓度下,杨树生物量会随着钙浓度的升高而增加,当氮的浓度为5mmol·L-1时最适钙浓度为5mmol·L-1;当氮浓度为20mmol·L-1时杨树幼苗的最适钙浓度为10mmol·L-1,其原因可能是因为钙的加入,使植物体内的氮钙比值趋向于更适宜的范围,低氮条件下适量的钙增加了植物体对氮素的吸收促进其生长,而高氮条件下适量的钙会提高植物体对氮的利用和转化减轻了氮浓度过高对植物体产生的毒害作用。所以适量的钙可以缓解氮过低或过高对植物生长产生的抑制,促进了植物的生长。
施加适量的钙可以促进植物的光合作用提高植物的净光合速率,积累植物的光合色素的产物,使叶片的净光合速率的增加、光合色素及光合产物累积等[25]。本试验表明,不施钙条件下不同氮处理的杨树幼苗叶片的净光合速率、气孔导度和蒸腾速率均维持在较低水平,而加入适量的钙后可提高杨树幼苗叶片的净光合速率、气孔导度和蒸腾速率,但当钙浓度过高时,会对杨树幼苗叶片的光合产生抑制作用,当钙浓度为20mmol·L-1时,不同氮浓度下的杨树幼苗叶片的净光合速率、蒸腾速率和气孔导度均低于不施钙的处理。前人在对松树和水曲柳的研究上得到了相似的结论[26-27]。由此可以得出结论:不同的钙浓度可以缓解氮过低或过高的供给时对植物带来的胁迫,其原因可能是因为钙促进了植物体内氮的吸收和转化,防止氮带来的抑制[28];其次可能是因为氮钙协同维持了植物体内的营养元素的平衡,保证可溶性蛋白及抗氧化酶的活性[29]以及光合作用的进行;最后钙离子的供应维持了叶片中叶绿体的完整性和稳定性,增强了光合代谢中的酶的活性,如二磷酸核酮糖羧化酶和PEP羧化酶,从而维持了较高的光合水平[30]。
当外界环境发生变化时,植物体受到胁迫时PSⅡ会发生相应的变化,呈现出显著下降的趋势,因此叶绿素荧光参数的变化经常被用来反映外界对植物体的胁迫程度。正常条件下植物的最大光化学效率应在0.80~0.84,当PSⅡ潜在活性中心受损时,植物的潜在光化学效率(Fv/F0)和最大光化学效率(Fv/Fm)均会有所下降,干扰光合电子传递、抑制光合作用的进行[31-32]。本试验结果表明,在不同氮水平下,杨树叶片的最大光化学效率(Fv/Fm)随着钙水平的提高表现为先升高后降低的趋势,表明氮钙协同可增加光合电子传递、提高光化学速率。设施油桃的研究与本试验结果相似,在两种不同的氮浓度下,设施油桃叶片的Fv/Fm表现为先升高后降低的趋势,说明在一定范围内,氮钙协同可以提高PSⅡ活性[17]。可能原因如下,一是适量Ca2+的可以维持植物体内各种营养元素的均衡,不会产生缺素症,提高植物体内抗氧化酶活性,保证杨树幼苗能够进行稳定的光合作用[33];二是Ca2+的施入促进了杨树的氮代谢,避免了叶片吸收的光能过剩不能全部进行转化与利用,光能过剩则会对光合器官造成不可逆伤害。
当氮处理相同时,杨树幼苗生长存在最适的钙浓度,使其生长、净光合速率、蒸腾速率、气孔导度、潜在光化学活性和最大光化学活性达到最大值;氮钙协同会促进杨树的生长发育,且表现为高氮处理条件下杨树幼苗生长适宜的钙浓度较高,低氮处理条件下适宜生长的钙浓度较低,整体表现为“高氮高钙,低氮低钙”的氮钙协同特征。因此在实际生产中要注意氮钙平衡,保证杨树正常生长发育。
