新疆特色奶疙瘩的菌群分析与抗衰老研究
2021-08-11李墨钊热米拉阿扎提郑晓冬周文文
李墨钊,热米拉·阿扎提,郑晓冬,周文文*
(1浙江大学生物系统工程与食品科学学院 杭州 310058 2华中农业大学生命科学技术学院 武汉 430070)
新疆奶疙瘩是一类历史悠久的结晶状发酵酸奶,风味独特,营养价值丰富,可加速人体代谢,提升人体抵抗力及提供维生素和矿物质,成为深受哈萨克族和维吾尔族等少数民族喜爱的传统食品。乳酸菌作为食品发酵中的重要微生物,是乳制品生产过程中的优势菌种。作者对新疆奶疙瘩开展研究,从中分离出多种乳酸菌,通过特定生化指标的测定筛选出有益生特性的优质乳酸菌资源[1]。课题组阿扎提等[2]对奶疙瘩中相关菌株进行分离纯化,筛选获得鼠李糖乳杆菌(Lactobacillus rhamnosus)R4 和瑞士乳杆菌(Lactobacillus helveticus)S4 等代表性菌株。秀丽隐杆线虫具有生命周期短,易于培养和与哺乳动物的高相似性等特征,可作为益生菌动物模型建立的良好材料,一直被认为是人体衰老领域研究的首选模式生物[3]。
本试验选用奶疙瘩样品,对其含有的益生菌进行高通量测序,研究其菌群多样性,确定优势菌群。前期对不同益生菌种的耐酸耐胆盐、疏水性、抑菌性及抗氧化等生化特性进行测定,从中分离筛选得到具有良好益生特性的菌株鼠李糖乳杆菌R4。本文以秀丽隐杆线虫为研究模型,研究R4 对其正常生理、生化指标(线虫寿命、产卵量、运动能力、热应激和氧化应激抵抗能力)的影响,探究R4的抗衰老作用,通过转录组测序分析其延寿机制,为进一步开发高效微生态制剂奠定基础。
1 材料与方法
1.1 试验材料
1.1.1 原料与试剂 奶疙瘩样品来自新疆维吾尔族自治区5 个地区(乌鲁木齐市、伊犁市、塔城市、阿泰勒地区和石河子南山地区) 的牧民家庭(图1),制作工艺采取家庭式传统方法发酵,输送途中用冰袋保藏,在实验室中4 ℃保存。

图1 来源于新疆不同区域的5 个奶疙瘩样品Fig.1 Five samples of milk knots from different regions in Xinjiang
琼脂糖、蛋白酶K、MgCl2、Platinum-DNApolymerase、dNTP-mix、PCR Buffer I、Chelex 裂解液、脱脂乳、M9 缓冲溶液、线虫冻存缓冲液、S medium、同期化漂白液、NGM 培养基、LB 培养基,杭州索莱尔博奥生物技术有限公司。鼠李糖乳杆菌R4(L.rhamnosus R4),本实验室分离保存。大肠杆菌(E.coli )OP50 和野生型秀丽隐杆线虫(C.elegans Bristol)N2 株,浙江大学动物科学学院杜爱芳教授惠赠。
1.1.2 主要设备与仪器 MLS-3750 型全自动灭菌锅,日本Sanvo 公司;电子天平,德国Sartotious公司;AF-100 型离心机,中国湘仪集团;Research plus 8 道移液器,德国Eppendorf 公司;WB-14 恒温水浴锅,德国Memmert 公司;Spectra mas Plus 384 型酶标仪,美国MD 公司;AF-100 型制冰机,上海Scotsman 公司;HiSeq 2000 型Illumina 测序仪,美国Illumina 公司;PTC-225Thermal PCR 仪,美国伯乐(Bio-Rad) 公司;StepOne 型定量PCR仪,美国应用生物系统(ABI)公司;LRH-250 型培养箱,上海一恒科技有限公司;U-RFL-T 型光学显微镜,日本Olympus 公司。
1.2 试验方法
1.2.1 基因组DNA 的提取 采用SDS-CTAB 法提取样品中菌种宏基因组DNA,通过紫外分光光度法测定其浓度,琼脂糖凝胶电泳判断其分子质量,加入无菌水稀释至样品质量浓度为1 ng/μL。
1.2.2 16 S rRNA V4 区、18S rRNA V4 及V9 区的PCR 扩增及高通量测序 以稀释后的基因组DNA 为模板,对16S rRNA V4 区、18S rRNA V4及V9 区进行PCR 扩增。引物使用带Barcode 的特异引物,对应区域:ITS1 区引物是ITS1-5FITS2;ITS2 区引物是ITS2-3F-ITS2-4;16S V4 区引物是515F-806R;18S V4 区引物是528F-706R;18S V9 区引物是1380F-1510R。使用PCR反应预混液Phusion®High-Fidelity PCR Master Mix with GC Buffer(New England Biolabs 公司)扩增。PCR 过程中使用高效酶,以保证反应的扩增效率和准确性。
按照高通量测序标准流程进行测试,构建PCR 产物文库,通过DNA 测序仪IUumina HiSeq 2000 对其进行双末端测序。为确保reads 的高质量,去除低复杂度、低质量序列及接头序列,得到clean reads,通过Overlap 拼接软件SOAP denovoV.1.0(华大自主产权)进行Overlap 拼接[4]。加入两端PCR 引物并确保引物3’端保守,处理后得到拼接后一致的序列,形成tag 序列并通过Mothur软件统计其丰度[5]。
1.2.3 线虫的同期化 秀丽隐杆线虫在NGM 培养基经72 h 以上的培育,部分转移成熟虫体至新平板开始培育、产卵。用5 mL M9 缓冲液洗至离心管中,1200 r/min 离心3 min,倒去上清。加入5 mL 新配的同期化漂白液,激烈振荡3 min,再次离心2 min,倒去上清。重复第1 步离心操作3 次,保留最后一次上清液2 mL,摇匀后转入不含大肠杆菌OP50 的平板中,20 ℃静置过夜后接种至NGM培养基。20 ℃培养20 d,此时线虫受精卵发育至L4 期,同期化完成。
1.2.4 线虫寿命及产卵、运动、氧化应激和热应激等表观性状 将线虫分为对照组和给药组。OP50为对照组,鼠李糖乳杆菌(L.rhamnosus)R4 为实验组,每组100 条。每天将其转移到新板上,20 ℃培养。当成虫不再产卵后,每2 d 挑取线虫1 次,直到线虫全部死亡时停止,使用软件SPSS 16.0分析数据[6]。重复寿命实验3 次,绘制图表。
将等数目同期化的对照组和实验组线虫挑取到各自平板中,每个平板1 条,产卵后以12 h 为周期转移,直至线虫不再具有生殖能力。培养所有留有线虫的平板,统计虫卵孵化至L4 期的成虫数量,即待测线虫的产卵量。
随机选取对照组和实验组中的线虫,以4 d为1 个周期培养,以其1 min 内运动的次数作为摆动频率的计数依据,分别记录2 组线虫的摆动频率数据,观测其运动能力。
将同期化至L4 的线虫分为对照组和实验组,每组有30 条,分别喂食大肠杆菌OP50 和乳酸菌,3 d 后将其转入溶液浓度为20 mmol/L 的过氧化氢中静置2 h,随后将线虫挑到NGM 培养基上,将培养基放在培养箱内20 ℃处理16 h,每小时统计线虫存活率,测定抗氧化应激能力[7]。重复实验3 次。
同期化对照组和实验组线虫至产卵期,各取出30 条转移至NGM 培养基(含OP50)中35 ℃热敷16 h 以上,统计其存活情况,分析线虫抗热激能力。当线虫对机械性刺激不再产生反应时,记录为已死亡[8]。
线虫成熟后再持续培养48 h,实验组加入乳酸菌,对照组加入大肠杆菌OP50。2 组线虫使用M9 缓冲液洗涤,冷冻、离心、冰浴破碎,提取上清液。SOD 酶活、CAT 酶活、MDA 含量用对应 的SOD、CAT 和MDA 检测试剂盒测定。
1.2.5 转录组重测序与实时定量PCR 分析 对提取的线虫样本RNA 使用DNA 聚合酶I 处理DNA 后用含有Oligo(dT)的磁珠对其mRNA 进行富集;添加打断试剂,将mRNA 打断成许多短片段,以这些短片段为模板合成一链cDNA;随后配制相应反应体系以合成二链cDNA,经纯化回收,修复黏性末端,cDNA 的3’端添加腺嘌呤“A”,连接接头,筛选片段大小,进行PCR 扩增反应,构建cDNA 文库;对构建好的文库进行质检和测序[9]。将过滤后的数据比对到参考基因组上,得到新转录版本,将其加入参考基因序列获得完整的参考序列,计算基因表达水平。参考KEGG 注释情况和官方分类,对差异基因在代谢通路上进行分类,聚集表达相近的基因成类,用R 软件phyper 函数富集分析差异基因,研究其作用及其所在信号通路。
对提取的线虫总RNA 进行反转录获得cDNA,对照已知秀丽隐杆线虫的衰老相关基因,对照Wormbase 及GenBank 中的序列用Primer 软件设计引物,按照荧光定量PCR 反应预混液Platinum SYBR Green qPCR Super Mix-UDG with ROX(Invitrogen 公司)试剂盒的方法进行实时定量PCR。
1.2.6 数据处理 实验重复3 次,取平均值,使用SPSS 16.0 软件,通过方差分析进行统计学检验。全部实验数据表示为±s,当P<0.05 时具统计学意义。
2 结果与分析
2.1 菌相分析
2.1.1 样品细菌总PCR 扩增结果 5 个样品细菌总DNA 经PCR 扩增,进行2%琼脂糖凝胶电泳,结果见图2。其中RS1.1、RS1.2、RS1.3、RS1.4 和RS1.5 分别由条带4,5,6,16 及17 表示,5 个样品的条带均清晰明亮。测定样品的密度分布大于50 ng/μL,总量大于100 μL,说明样品符合测试要求,可进一步实验。

图2 16S rDNA V6 区PCR 扩增产物电泳图Fig.2 PCR products of V6 region in 16S rDNA
2.1.2 高通量测序结果分析 5 个奶疙瘩样品经测序得到3.10 Gb 原始数据,对5 个样品的全部Effective Tags 序列聚类成为OTUs,并对tags 序列去冗余,选出unique tags 序列[10]。根据物种注释特征,对比这5 个样品序列,比较16S rDNA V6 区域的初始数据和获得的tags 结果,发现绝大多数选出的unique tags 序列处于“属”这一分类单元。选出最大相对丰度前10 名的属对应OTUs 的系统发生关系的数据和对应OTUs 的相对丰度信息(图3)。测序结果表明奶疙瘩样品中的链球菌属(Streptococcus)、假单胞属(Pseudomonas)、乳球菌属(Lactococcus)、乳杆菌属(Lactobacillus)为其优势菌群。其中,在除RS1.1 样品外的4 个样品中乳酸菌属均为优势菌,即:乳酸菌在RS1.2 样品中有34.10%,在RS1.3 中有17.93%,在RS1.4 中达54.53%,在RS1.5 中也达40.05%。对比5 个样品菌群含量,推测RS1.1 中高含量的假单胞菌属可能是在家庭作坊生产时被污染产生的。

图3 OTUs 丰度聚类图Fig.3 Operational taxonomic units abundance clustering diagram
2.2 代表性乳酸菌R4 的性质及对线虫生理、生化指标的影响
2.2.1 鼠李糖乳杆菌R4 的性质 前期研究[2]表明,鼠李糖乳杆菌R4 为样品中乳杆菌属中功效优良的益生菌。在pH 3.0 和0.3%胆盐浓度条件下分别处理3 h 后,鼠李糖乳杆菌R4 分别具有87.1%和74.9%的存活率,最高的抗氧化活性,对胆固醇和甘油三酯降解率高达50.97%[11]。基于其优良性质对R4 饲喂的线虫在不同条件下实验,进一步探究R4 的功效。
2.2.2 鼠李糖乳杆菌R4 对线虫产卵量和运动能力的影响 线虫的产卵量和运动能力可作为线虫寿命的相关指标,用于全面评价乳酸菌对其的抗衰老作用。产卵为线虫机体生长发育的有效生物指标之一。实验中,将秀丽线虫分别放在涂有R4和大肠杆菌OP50 两种菌株的已灭活组、活菌和上清液的NGM 培养基上培养,60 h 后分别计数各培养基中线虫的产卵数(图4)。饲喂R4 活菌的实验组线虫产卵数量为347.2±4.53,而饲喂OP50 活菌的对照组的产卵量为315.4±8.99。饲喂灭活R4 菌株的实验组产卵量为323.5±3.61,对照组的产卵量为306.2±5.90。上述结果表明,R4 菌株的3 种处理方式均对线虫的生殖系统没有产生明显影响,反而有一定的促进作用。

图4 鼠李糖乳杆菌R4 对线虫产卵量的影响Fig.4 Effect of L.rhamnosus R4 on nematodes brood sizes
线虫的肌肉功能强、弱可由其运动状态测定,其运动能力随年龄增长而下降,于是将8日龄秀丽线虫的寿命作为指标,探究R4 对其运动能力的影响[11]。线虫运动轨迹见图5,身体摆动频率的测定结果显示:实验组中喂食R4 的线虫在20 s 内身体摆动频率为11.73±1.82,均大于对照组(OP50)11.35±0.91,然而无显著性差异。在第8~12天内,与对照组相比,实验组秀丽线虫的运动能力和活跃程度显著增强,死亡数量显著降低(图6)。运动能力的减弱会导致线虫需要的能量减少和呼吸水平降低,进而氧化损伤减少,这说明R4 延缓线虫衰老。

图5 线虫的运动轨迹(放大为25 倍)Fig.5 The actions of C.elegans(at 25×magnification)

图6 鼠李糖乳杆菌R4 对线虫运动能力的影响Fig.6 Effect of L.rhamnosus R4 on the movements of C.elegans
2.2.3 鼠李糖乳杆菌R4 对线虫寿命的影响 前面研究表明,鼠李糖乳杆菌R4 与大肠杆菌OP50饲喂组相比,R4 饲喂蠕虫的平均寿命增加,R4供体虫的寿命为(31.05±0.54)d,比对照虫的寿命(22.81±0.62)d 长36.1%[2]。本研究对反映线虫存活能力的重要指标——热激寿命和氧化应激寿命进行探讨。在抗热激实验中,与对照组的(8.34±1.51)h 相比,R4 明显延长了线虫的平均寿命至(12.55±0.68)h。线虫对过氧化氢导致的氧化损伤的耐受能力也明显升高39.9%,其平均寿命达(9.78±1.86)h,延缓寿命趋势明显(图7)。在压力条件下,鼠李糖乳杆菌R4 对线虫的抗氧化应激能力的显著提高可能是其对延缓线虫寿命的机制之一[12],这说明在不良环境下,奶疙瘩中乳酸菌对线虫有一定的保护作用。

图7 不同条件下鼠李糖乳杆菌R4 对线虫平均寿命的影响Fig.7 Effects of different conditions on the lifespan of C.elegans with L.rhamnosus R4
2.2.4 鼠李糖乳杆菌R4 对线虫生化指标的影响SOD 与CAT 的表达同线虫抵御环境氧化应激相关。用R4 处理后的秀丽线虫体内SOD 与CAT等抗氧化酶的活力显著提高(图8),这印证了R4能够增强线虫抵御环境氧化应激能力的结果,说明其通过提高线虫体内SOD 和CAT 等的酶活来提高其抗氧化应激水平。对比对照组CAT 酶活(33.57±4.52)U/mg,R4 活菌处理组中CAT 酶活明显提高到(47.43±0.85)U/mg;上清液处理组中CAT 酶活(23.01±2.87)U/mg 虽比对照组OP50(25.63±3.46)U/mg 稍有减少,但差异不显著。而R4 灭活组CAT 酶活与OP50 对照组相比大幅降低,表明R4 活菌的CAT 酶活力最强。从图8可以发现,喂食R4 活菌、上清液和灭活菌体等3 种处理组的线虫体内MDA 含量均显著降低,表明鼠李糖乳杆菌R4 具有抗脂质过氧化的作用。
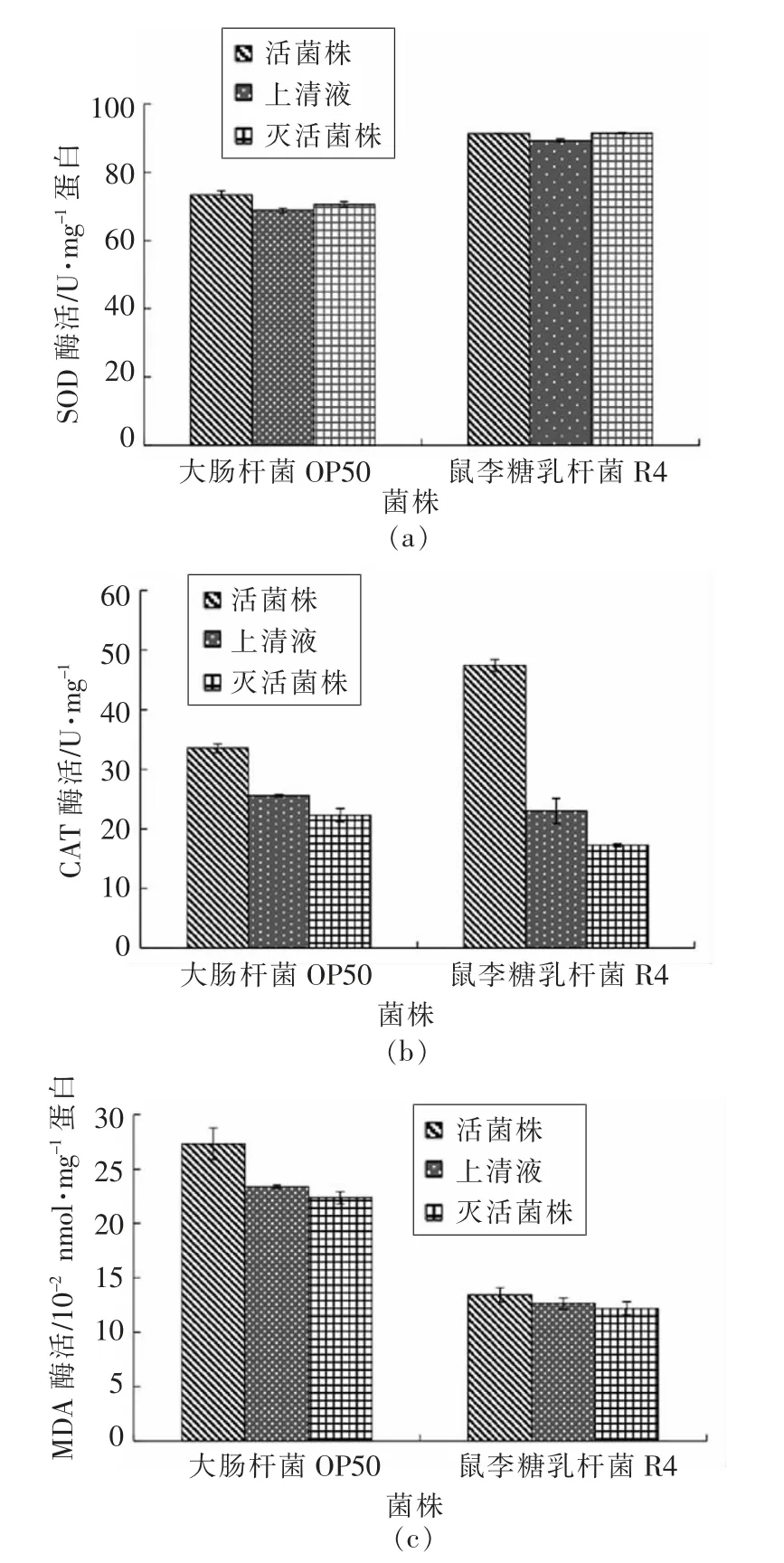
图8 鼠李糖乳杆菌R4 对线虫体内生化指标的影响Fig.8 Effects of L.rhamnosus R4 on antioxidative enzymes of C.elegans
2.3 转录组与基因转录分析
转录组测序显示,饲喂鼠李糖乳杆菌R4 的线虫体内出现大量与编码寿命相关蛋白的基因和次级代谢通路基因转录变化。这其中影响线虫寿命的蛋白为乙酰辅酶A 脱氢酶(acdh-1)、卵黄原蛋白(vit-4)、脂肪酸脱氢酶(fat-7)和脂肪酸结合蛋白等。同时,线虫在一定浓度的R4 影响下氧化还原,与物质代谢的相关基因也受到影响,主要有:过氧化氢酶(ctl-1)、3-羟酰-辅酶A 脱氢酶(hacd-1)、超氧化物歧化酶(sod-3)、饮食限制下调蛋白(drd-1)、水化酶(ech-7)以及脂肪酸结合蛋白(fat-5)。此外,秀丽线虫体内daf-16 基因的表达水平出现明显上调(6.12 倍),其在胰岛素信号通路中起关键作用。鼠李糖乳杆菌R4 能抑制水化酶和乙酰辅酶A 脱氢酶(Enoyl-CoA Hydratase和Acyl CoA Dehydrogenase) 等合成基因的表达(E 值均为0)。
随机对线虫体内与抗衰老有关的基因进行RT-PCR 测试,以增强测序结果的可信性,发现显著影响的6 个基因,如图9所示。在氧化应激条件下,以管家基因gpd-1 为内参,用R4 处理的线虫体内sod-3、hsp-16.2、ctl-1 和daf-16 基因的mRNA 水平均明显升高。该实验表明在氧化应激条件下,R4 在激活线虫表达sod-3 的同时,也显著上调热激蛋白hsp-16.2 的表达量。sod-3 基因编码的蛋白具有清除生物体内过量的超氧化物自由基的功能,可以阻止细胞的氧化损伤;热激蛋白在生长温度过高时帮助新合成的肽链正确折叠、伸展以发挥正常功能,保持生物活性,防止蛋白质变性,有效避免细胞受到热损伤。研究结果表明:在氧化应激条件下,鼠李糖乳杆菌R4 可通过daf-16转录因子的激活促进下游sod-3 基因的激活,使细胞获得氧化应激保护。

图9 鼠李糖乳杆菌R4 对线虫体内与衰老相关基因转录水平的影响Fig.9 Influences of L.rhamnosus R4 on the mRNA expression of ageing-associated genes in C.elegans
daf-2 基因主要负责线虫体内寿命延长和发育停滞的调控,与能量摄取紧密联系,被认为作用于胰岛素信号通路的下游抑制daf-16 表达[13]。然而,实验结果显示,daf-2 对daf-16 没有产生抑制作用,说明R4 并非直接抑制daf-2 来激活daf-16,同时说明R4 对daf-2 的影响较小,对daf-16的影响较大。由此可推断鼠李糖乳杆菌R4 对线虫寿命的调控可能依赖于daf-16/FOXO 信号通路,通过激活通路下游抵抗压力的基因来清除自由基,从而减慢衰老。经R4 处理后,秀丽线虫体内sir-2.1 与daf-2 基因mRNA 的表达水平的明显下调,说明鼠李糖乳杆菌R4 或许通过Insulin/IGF-1通路参与线虫寿命的正向调控,延缓衰老。
Insulin/IGF-1 信号转导通路上的sod-3 转录水平的提高同样证明R4 可以激活下游转录因子daf-16。影响线虫发育的“生物钟”基因ctl-1 是daf-16 的下游基因,其表达量的上调也进一步证实R4 激活了daf-16 的转录活性。本研究表明鼠李糖乳杆菌R4 对线虫的正向保护作用依赖于daf-16/FOXO 通路。此外,本文推测R4 的作用可能是通过曲线调节,即以间接的方式实现调控。当氧化压力较大时,R4 通过调节sir-2.1 基因的表达,从而进一步调控寿命。通常情况下R4 对寿命的正调控多被认为是通过sir-2.1 实现的。
3 结论
本研究对新疆奶疙瘩进行菌群分析,对其中的高活性益生菌鼠李糖乳杆菌R4,以秀丽隐杆线虫为模型,测试其对线虫寿命及其它生理、生化指标的影响,结合转录组重测序等手段揭示其对线虫寿命的调节方式。R4 处理的秀丽线虫的产卵率和运动频率上升,对热应激和氧化应激胁迫的抵抗力有所提高,同时线虫体内SOD 和CAT 酶活显著升高,而MDA 的含量下降。可见R4 具有较强的延寿作用。实验显示:该菌株的活菌不仅可以延缓线虫寿命,还能够保护线虫在不良环境下免受机体损伤。对寿命和衰老相关基因的监测表明,乳酸菌R4 主要影响daf-16、daf-2、sir-2.1、ctl-1、hsp-16.2 和sod-3,R4 主要通过激活胰岛素信号通路(Insulin/IGF pathway)下游的daf-16 基因增强抗应激能力和延长寿命。R4 活菌处理组对秀丽线虫sod-3 mRNA 水平的上调表明其对daf-16转录因子的激活作用。而daf-16 的下游基因ctl-1表达量的上调也证实鼠李糖乳杆菌R4 活菌激活了daf-16 的转录活性。
