‘瑞雪’苹果脱毒幼苗接穗采集区的研究
2021-08-10冯瑞芳闫雷玉孙建春孙鲁龙赵政阳
冯瑞芳,闫雷玉,孙建春,孙鲁龙,赵政阳
(西北农林科技大学 园艺学院,陕西杨凌 712100)
苹果树体感染病毒是降低果实品质的重要因素之一[1]。苹果树体感染病毒后终生带毒且无有效的防治措施,感染病毒后的树体果实品质变劣,单株产量降低[2-3],感病严重时还会导致树体整株死亡[4-5]。控制病毒病害蔓延最有效的措施是栽植脱毒苗木。‘瑞雪’是西北农林科技大学选育的黄色晚熟苹果优质新品种[6],具有丰产、大果、味美等特点,是晚熟品种中少有的黄色品种[7]。‘瑞雪’苹果新品种推广过程中脱毒苗木的繁育非常重要,芽接技术可以提高接穗利用率、扩大繁殖系数以及加快‘瑞雪’苹果的推广速度。此外,从幼龄脱毒苗上采集接穗降低嫁接苗感染病毒几率的同时还可实现采穗植株密植化,达到高效率苗木生产的目标。由于芽的异质性导致不同部位的芽体质量不同,使得苗木嫁接后表现也不同。核桃嫁接的研究表明,1a生枝条中部芽的成活率较高,新梢长到30~50cm掐头后接穗的利用率高[8-9]。‘光辉’海棠和‘红肉’苹果芽接技术中发现,选择枝条中部的芽成活率显著高于其他部位的芽[10]。综上所述,单芽接穗的选择对于‘瑞雪’嫁接繁殖具有重要的意义。
目前,关于接穗的选择主要集中在核桃的研究中,关于如何从脱毒苹果幼树上选择接穗的研究较少。由于从脱毒幼树上采集接穗有芽体营养不足、分化较差的风险,因此,有必要对不同节位的芽、叶以及采集接穗嫁接后的嫁接苗进行分析。本研究选择长势一致的1a生和2a生幼龄‘瑞雪’脱毒自根苗为试验材料,从树体基部向上每10个芽为一个采穗区,共分为5个采穗,对照为带毒母树1a生枝条中部的采穗区,采集各采穗区内1a生枝上的叶片及单芽,采集的叶片用以比较叶面积、叶片厚度、叶片栅栏组织厚度、叶绿素、淀粉、可溶性糖、可溶性蛋白、多胺等生理指标的差异;采集的单芽嫁接至M26矮化自根砧上用以比较嫁接苗的长势。通过不同采穗区嫁接前叶片指标和嫁接后芽接苗长势的分析,筛选出最佳的接穗采集区,为‘瑞雪’幼龄脱毒苗单芽接穗的选择提供科学参考。
1 材料与方法
1.1 试验材料与设计
1.1.1 试验材料 试验地点位于陕西省白水县西北农林科技大学白水苹果试验站(109°16′~109°45′E, 35°4′~35°27′N),海拔860 m左右,地处黄土高原沟壑丘陵区,温带半湿润大陆性季风气候。
试验材料为1 a生和2 a生‘瑞雪’脱毒自根苗,脱毒自根苗来源于西北农林科技大学园艺学院实验室,运用茎尖超低温和热处理技术脱除病毒,经RT-PCR检测已脱除APSV、ASGV、ACLSV 3种潜隐性病毒,在2018年4月和2019年5月炼苗1周后,定植于西北农林科技大学白水苹果试验站,定植前进行土壤改良和土壤消毒;定植株距0.6 m,行距3 m;M26脱毒矮化自根砧木2018年4月定植于试验站,株距0.6 m,行距3 m。试验对照试材为带毒当年生枝条中部饱满节位。对试验所选试材进行常规幼苗管理措施,试材生长一致。
1.1.2 试验设计 1 a生树体从其基部向上每10个芽作为一个采穗区,2 a生树体基部50 cm向上每10个芽作为一个采穗区,如表1所示,将幼树分为5个采穗区,采集当年生枝条上的单芽和叶片,2019年9月1日选取1 a和2 a年生幼树长势一致且健康的幼苗各3棵,根据划分的采穗区,每个采穗区采集3颗单芽接穗,使用带木质嵌芽接方法嫁接到M26脱毒砧木,嫁接口高度距地面20 cm,对成活后的嫁接苗进行常规管理。嫁接时采集各采穗区的叶片洗净后液氮冻存备用,每个处理重复3次。
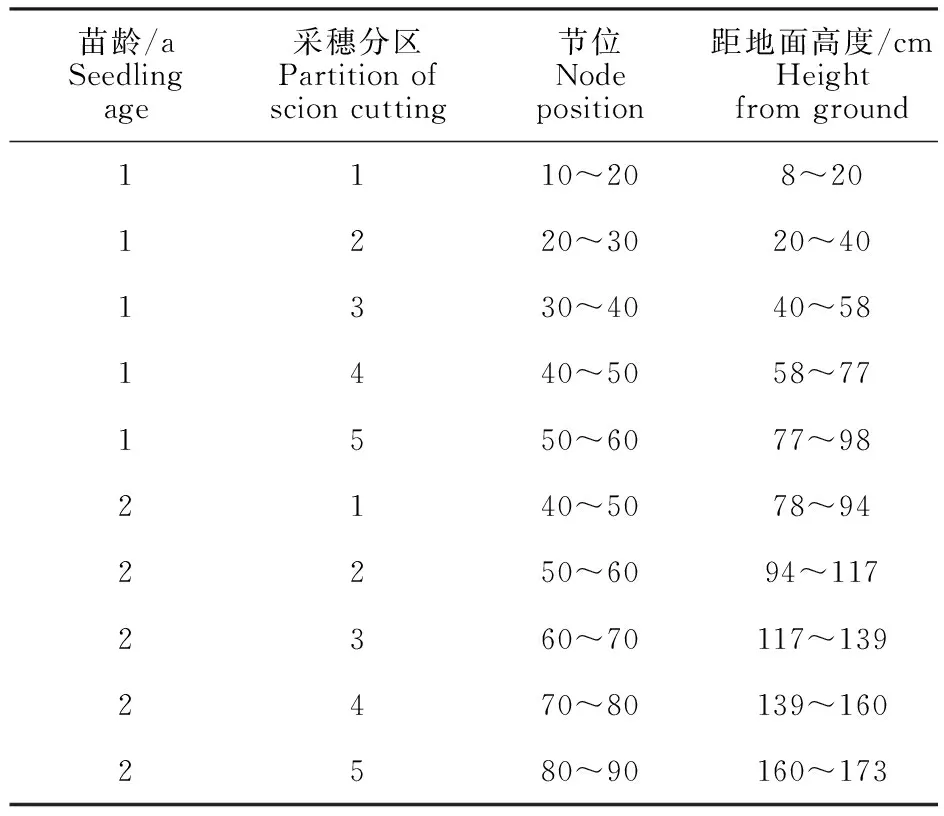
表1 ‘瑞雪’脱毒自根苗采穗分区Table 1 Partition of scion cutting of ‘Ruixue’ free-virus self-root seedlings
1.2 测定指标及方法
1.2.1 叶面积测定 运用托普云农YMJ-D型手持活体叶面积测量仪测得。
1.2.2 叶片解剖结构测定 将新鲜叶片避开叶脉剪成1 cm的方块,经固定、脱水、透明、浸蜡、包埋、切片、展片、脱蜡、番红固绿染色、封片后制成永久性切片;奥斯巴林正置荧光显微镜拍得照片,用奥斯巴林配套软件测得叶片厚度和栅栏组织厚度[11]。
1.2.3 叶片生理生化指标测定 叶绿素含量用丙酮法测定[12];淀粉含量用蒽酮比色法测定[13];可溶性糖含量用蒽酮比色法测得[13];可溶性蛋白质含量用考马斯亮蓝法测定[14];多胺含量测定方法,参照刘俊等[15]的高效液相色谱法测进行多胺测定,色谱条件为流动相:V(甲醇)∶V(乙腈)∶水=58∶2.5∶39.5,流速1 mL/min,波长230 nm,多胺标准品购于上海源叶生物有限公司官网,多胺测定仪器使用日本岛津LC-30A超高效液相色谱仪。
1.2.4 脱毒嫁接苗树高、干径测定 2020年10月初嫁接苗趋于停止状态测定嫁接苗树高和干径,用卷尺由嫁接口处至最高生长点测定幼苗的高度,干径取嫁接口上10 cm光滑部位,用游标卡尺在东、西、南、北方向各测量一次后取平均值。
1.3 数据分析
利用SPSS 19.0软件进行方差分析,多重比较采用LSD 法(P<0.05),Excel 2013软件进行表格和图表制作。
2 结果与分析
2.1 不同采穗区叶面积的变化
叶面积测定结果表明,1 a生‘瑞雪’(图1-A)随着节位增高叶面积有增大的趋势,1 a生第3采穗区叶面积最大,为5 841.76 mm2,叶面积最小的是第5采穗区。2 a生‘瑞雪’叶面积(图1-B)随着节位增高叶面积有先增后降的趋势,第3采穗区的叶面积最大,为6 181.81 mm2,高于对照4.3%,叶面积最低的是第1采穗区。
2.2 不同采穗区叶片解剖结构
叶片结构的变化与叶片质量正相关,影响叶片活性和光合作用,叶片通过光合作用产生的有机物为腋芽提供充足的营养物质[16-17]。叶片厚度的结果表明,随着节位升高叶片厚度呈先升高后降低的趋势,1 a生和2 a生‘瑞雪’在第3采穗区叶片厚度分别为242.39 μm和244.07 μm;1 a生树体低于对照6.1%,2 a生树体与对照相比没有显著性差异(图2)。
叶片栅栏组织厚度(图3),1 a生和2 a生‘瑞雪’叶片栅栏组织厚度随节位升高呈现先增加后降低趋势,1 a生和2 a生‘瑞雪’第3采穗区栅栏组织分别为101.55 μm和105.46 μm, 1 a生树体第3采穗区显著低于对照10.2%,2 a生‘瑞雪’第3采穗区的栅栏组织与对照无显著性差异。
2.3 不同采穗区生理生化指标的变化
2.3.1 叶绿素含量 叶绿素在光合作用对于能量捕获和传递至关重要,叶绿素的含量影响有机物质的积累[18]。图4中1 a生‘瑞雪’5个采穗区间叶绿素含量没有显著性差异并均低于对照。2 a生‘瑞雪’随着节位的升高叶绿素含量增加,叶绿素含量最少的是第1采穗区,第3采穗区叶绿素含量最高为3.96 mg/g,与对照在0.05水平上没有显著差异。
2.3.2 碳水化合物含量 碳水化合物的积累有利于腋芽的饱满程度,是芽分化的重要物质基础[19]。1 a生‘瑞雪’淀粉含量为2.12 mg/g,显著低于对照 20.8%(图5-A);2 a生‘瑞雪’的第3采穗区淀粉含量最高,为2.61 mg/g,显著低于对照2.9%(图5-B)。
1 a生‘瑞雪’可溶性糖含量呈先升高后降低趋势,第3采穗区可溶性糖含量最高,为2.25 mg/g,显著低于对照34.8%(图6-A)。2 a‘瑞雪’可溶性糖含量最高的是第3采穗区,含量为 3.36 mg/g,低于对照2.6%(图6-B)。
2.3.3 可溶性蛋白含量 蛋白质是细胞分裂、分化的主要物质基础, 其含量变化与花芽分化也有密切关系[20]。1 a生和2 a生‘瑞雪’5个采穗区中第3采穗区可溶性蛋白质含量最高,分别为 0.74%和0.73%,分别低于对照2.6%、3.9% (图7)。
2.3.4 多胺含量 多胺是细胞学中的重要调控物质,缺少多胺使细胞增殖分化困难,导致细胞生长缓慢。另外,多胺可显著促进果树花芽形成[21-22]。
1 a生和2 a生‘瑞雪’叶片中亚精胺(Spd)含量最高,在第3采穗区处到达高峰(47.75 μmol/g)之后下降,与对照无显著性差异(表2)。2 a生树体第3采穗区含量最高,为45.78 μmol/g,与对照无显著性差异。
1 a生‘瑞雪’5个采穗区中腐胺(Put)含量差异不显著,第3采穗区Put含量最高,为26.83 μmol/g,显著低于对照5.21%(表2)。2 a生‘瑞雪’第3采穗位Put含量为29.59 μmol/g,与对照相比在0.05水平上有显著性差异(表2)。

1 a生和2 a生‘瑞雪’叶片中精胺(Spm)是3种多胺中含量最低的,1 a生‘瑞雪’第3采穗区含量最高为21.44 μmol/g,低于对照14.37%。2 a生树体第3采穗区Spm含量显著高于其他采穗区,为22.56 μmol/g,与对照无显著差异。
2.4 不同采穗区接穗的嫁接苗生长量
如表3所示,选自1 a生自根苗第3采穗区的嫁接苗的生长量显著高于其他采穗区,树高高于对照16.9%,干径高于对照9.2%,总节数高于对照17.8%,侧枝数与对照无显著差异;选自2 a生自根苗第3采穗区的嫁接苗生长量高于其他4个采穗区,树高显著高于对照15.8%,干径高于对照8.7%,总节数高于对照9.6%,侧枝数高于对照6%。脱毒苗除1 a生第1采穗区的干径和2 a生第1采穗区嫁接苗的干径低于对照,其他采穗区芽接苗的树高、干径、侧枝数和总节数均高于对照。

3 讨 论
3.1 叶片质量对腋芽质量的影响
叶片质量可以体现树体的营养生长状态,叶面积大接受光照射面积也大,有利于制造有机物质[23]。研究表明叶片解剖结构与光合作用密切相关,光合速率与叶片厚度、叶片栅栏组织正相关,叶片栅栏组织厚度越厚越有利于光能捕捉,此外叶片也有一定储存有机物质的能力[24-25]。源-库学说理论中,树体的叶片为代谢源,同节位的芽为代谢库,代谢源的强度最先影响到离代谢源最近的代谢库[26]。本研究发现树体中部采穗区的叶片面积、叶片解剖结构测得结果优于基部和顶部,30~40节(1 a)、60~70节(2 a)叶片中淀粉、可溶性糖、可溶性蛋白质、多胺等内含物高于其他节位,此部分的叶片制造有机物质的能力高,此外,选用上述采穗区内芽作为接穗的芽接苗生长势优于其他节区,基于源-库学说可以明确地解释这一现象,处于此采穗区的腋芽内有机物质多、芽体饱满,可为翌年嫁接苗的生长提供必要的物质基础。
3.2 芽质量对嫁接的影响
芽的异质性导致同一枝条不同部位的芽存在着大小、饱满程度等一些差异[27]。秋季芽接应选取枝条中段充实饱满的芽作为接芽[28]。桑娟萍等[29]在核桃芽接的研究中发现,取同一枝条不同部位的芽子做接穗,中部芽子的嫁接成活率为78%,基部的芽子嫁接成活率为70%。赵辉等[30]在研究核桃不同部位芽对嫁接成活率的影响中发现中部芽子成活率最高,上部次之,且中部芽子较上部和下部的芽生长最快。这与本研究结果一致,本研究中1 a生幼苗的中上部和2 a生树体的中部叶片结构和生理生化指标则处于较高的水平,使得其腋芽饱满程度高于其他节位的腋芽。优质饱满的芽体有利于嫁接后成活率、愈合情况以及翌年的长势。
糖类作为植物主要的同化物质,在植物的生长发育和形态建成中起着重要的作用[31],研究表明花芽分化进度和蔗糖、可溶性糖的含量正相关[32]。多胺作为具有活性的物质,在植物成花转变中起调控作用[33]。Put、Spd有利于龙眼的营养生长,Spm有利于花芽分化[34]。奈李花芽分化期间Put含量降低,Spd、Spm出现高峰。Put是Spd和Spm合成的底物、Put含量直接影响到后两者的含量[35]。有研究表明, miR156过量表达抑制植物开花时期, ‘寒富’离体试管苗中miR156的表达量显著高于田间苗[36]。由于试管苗中miR156的表达量高于田间苗,由试管苗获得的脱毒自根苗可能会出现开花延迟等现象,那么接穗的选择不光要考虑芽体的质量还需考虑果树不同节位花芽分化的问题。
本研究中1 a生树体30~40节(第3采穗区)、2 a生树体60~70(第3采穗区)节的可溶性糖、可溶性蛋白质含量、多胺总含量显著高于其他采穗区;1 a生、2 a生树体各节区 Spd含量最高,Put含量次之,Spm含量最低。本研究发现1 a生‘瑞雪’30~40节(第3采穗区)和2 a生‘瑞雪’60~70节(第3采穗区)叶片中的叶面积、可溶性糖、淀粉、可溶性蛋白和多胺含量最高,使用此采穗区嫁接成活后苗木的营养生长优于其他节区并显著高于对照。
4 结 论
通过对1 a生和2 a生‘瑞雪’脱毒自根苗各个采穗区叶片生理指标的分析比较,综合不同采穗区芽接苗成活后生长势的分析,初步得出以下结论:生产中使用幼龄‘瑞雪’脱毒自根苗采集接穗时,接穗的最佳采集区域为:1 a生‘瑞雪’树体30~40节,对应树高40~58 cm;2 a生树体的 60~70节,对应树高117~139 cm。
