张家口籍汉族人CD14基因及其-159C/T位点多态性与心血管疾病关联研究
2021-08-08周璐瑾崔思瑶陈素艳李喜龙刘俊杰闫珂王瑾洁王文栋常晓彤
周璐瑾 崔思瑶 陈素艳 李喜龙 刘俊杰 闫珂 王瑾洁 王文栋 常晓彤
河北北方学院医学检验学院,张家口,075000,中国
心血管疾病(cardiovascular disease,CVD)通常由高血压、高血脂和动脉粥样硬化引发,在临床上主要表现为心悸、眩晕、紫绀、呼吸困难和卒中等症状[1],且通常伴有较多并发症,严重危害人类健康。我国CVD 发病率高,死亡率位居首位。随着我国人口老龄化与社会城镇化进程的发展,我国心血管疾病危险因素流行趋势明显[2],防治问题刻不容缓。
炎症反应对心血管疾病有显著影响[3-4]。单核细胞和巨噬细胞是最早被鉴定出与心血管疾病有关的免疫细胞,在动脉粥样斑块内含有少量的激活B 细胞和树突状细胞以及大量的单核巨噬细胞和激活T 淋巴细胞[5]。单核细胞具有明显的异质性,依据其表面分子白细胞分化抗原14(cluster of differentiation antigen 14,CD14)和CD16 的表达水平可划分为经典型(CD14++CD16-)、中间型(CD14++CD16+)和非经典型(CD14+CD16++)三种单核细胞nianfu 亚群,且三种单核细胞亚群可相互转化[6]。中间型单核细胞亚群参与了脑缺血性中风和心脏栓塞等心血管疾病的发病过程[7],在预后不良冠心病患者体内含量显著升高,有望作为治疗冠心病的靶细胞[8-9]。单核细胞表面CD14 蛋白含量的变化机制对各单核细胞亚群之间的转化具有极其重要的意义[10]。
CD14 基 因位于人类5 号染色 体5q23-31 区,其编码产物蛋白具有2 种形式,一种是膜结合型CD14(membrane-bound CD14,mCD14),通过糖基磷脂酰肌醇(glycosylphosphatidylinositol,nian’fuGPI)锚定结合于单核细胞、巨噬细胞和中性白细胞的膜外侧;另一种是可溶型CD14(soluble CD14,sCD14),游离在血浆中[11]。CD14 蛋白可作为许多微生物产物受体,与相应配体结合激活单核细胞、巨噬细胞和血管内皮细胞等,分泌细胞因子及生长因子,调控黏附因子的表达,加速蛋白质糖化产物的形成并导致其在血管内堆积,引起血管平滑肌细胞的增生和动脉内中膜增厚[12]。由此可见,CD14 在炎症、动脉粥样硬化的发生过程中起着重要作用,但其在心血管疾病中的分子遗传机制尚不清楚。已知CD14 基因存在-159C/T(rs2569190)多态性位点,TT 纯合子的sCD14 水平明显高于CC 和CT 基因型的携带者,且干扰素-γ(interferon-γ,IFN-γ)和白介素4(interleukin 4,IL-4)反应分别与血清sCD14 水平呈正相关和负相关[13]。本研究利用生物信息学方法、聚合酶链反应-限制性片段长度多态性(polymerase chain reaction-restriction fragment length polymorphism,PCRRFLP)和测序等技术对CD14 基因及其编码产物进行分析,研究CD14 基因-159C/T 位点多态性与心血管疾病的关系,从而探讨CVD 的分子遗传机制。
1 资料与方法
1.1 资料及其来源
收集河北北方学院附属第一医院心血管疾病患者血浆标本75 例,其中男性患者42 例,女性患者33例;健康对照组125 例,包括男性患者80 例,女性患者45 例,每人采静脉血5 mL。本研究经医学伦理委员会同意,且所有纳入者均知情同意。从Uniport 数据库(https://www.uniprot.org)查询获得人CD14_HUMAN(P08571)蛋白序列[14]。
1.2 实验方法
1.2.1 DNA 提取及目的片段扩增
根据全血基因组DNA 提取试剂盒说明书,采用SMA4000UV-Vis 超微量紫外分光光度计测定仪测定DNA含量与纯度。根据测得的DNA 浓度配制50 μL 的PCR 体系(10×PCR Buffer 10 mmol·L-15 μL;dNTP 2.5 mmol·L-14 μL;上游引 物5′-ATCATCCTTTTCCCACACC-3′10 μmol·L-13 μL;下游引 物5′-AACTCTTCGG CTGCCTCT-3′ 10 μmol·L-13 μL;模 板DNA,浓 度为400 ng/DNA;200 μmol·L-1的Tap 酶 0.25 μL;最后蒸馏水补足至50 μL),进行基因扩增(95 度预变性10 min,94 度变性40 s,61 度退火40 s,72 度延伸1 min;重复35 个循环,72 度补平10 min,4 ℃保存),产物经0.8%琼脂糖凝胶电泳,结果通过Oemge 紫外投射仪进行检验,理论目标条带为296 bp。
1.2.2 产物酶切及鉴定
根据PCR 产物,进行酶切。HaeⅢ酶识别和切割位点序列为GGCC,若位点序列改变,则无法切割。HaeⅢ酶37 ℃下反应3 h 后,产物经3.5%琼脂糖凝胶电泳,紫外投射仪检验,根据所得结果进行分型。理论目标条带:CC 型为152 bp 和141 bp 两条带;CT 型为296 bp、152 bp 和141 bp 三条带,TT 型为296 bp 一条带。将PCR 反应产物进行测序,比对测序结果,确定最终分型。
1.3 统计学方法
应用SPSS23.0 统计软件对正常对照组和CVD患病组的CD14 基因型对比情况进行卡方检验统计,其中,年龄分布的计量资料采用独立样本t检验。P<0.05,表示差异有统计学意义。
1.4 生物信息学分析
1.4.1 人CD14 蛋白理化性质与结构分析
利用生物信息学分析工具(Tab.1)分析CD14 基因分析蛋白的理化性质、亲疏水性、信号肽、亚细胞定位,预测其二级结构与三级结构,并进行质量评估。

Tab.1 Bioinformatics analysis tools
1.4.2 人CD14 蛋白互作分析及基因功能注释
使 用STRING 数据库(https://string-db.org/)[15]挖掘以人CD14 蛋白为中心结点的蛋白互作网络,采用Cytoscape 软件v3.6.1 版本[16]完成可视化,筛选出关联基因。然后使用DAVID 数据库(Visualization,Annotation and Integrated Analysis,https://david.ncifcrf.gov)[17]对CD14 和关联基因进行富集分析,找出显著富集的生物学功能注释(P≤ 0.05 和 Count ≥4 genes)[18]。
2 结果
2.1 酶切检测基因型结果
PCR 产物酶切后,电泳结果如Fig.1A、B 所示,测序鉴定结果见Fig.1C、D、E,据此区分三种不同的基因型。
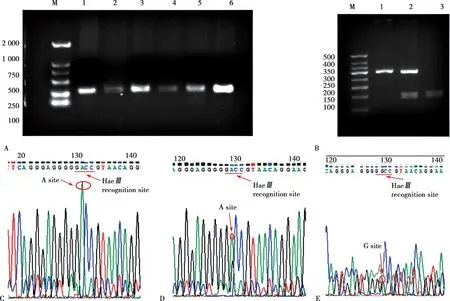
Fig.1 CD14 gene PCR product sequencing results and enzyme digestion results
基因型分布频率见Tab.2,等位基因分布频率见Tab.3。用χ2检验分析患病组和正常对照组等位基因频率及基因型有无统计学差异以及是否符合Hardy-Weinberg 平 衡。对照组 和患病 组CD14-159C/T(rs2569190)基因多态性分布频率符合 Hardy-Weinberg平衡。Tab.2 中,TT 基因型分布频率在患病组(56%)和正常对照组(40.8%)之间有显著统计学差异(P<0.05)。Tab.3 显示,对照组和患病组之间等位基因频率也存在显著差异(P<0.05)。结果提示,心血管疾病与CD14-159C/T(rs2569190)等位基因有关联。

Tab.2 CD14 genotype comparison between normal control group and diseased group

Tab.3 Comparison of CD14 allele frequency between normal control group and diseased group
2.2 CD14 蛋白的生物信息分析
2.2.1 理化性质分析结果
CD14 基因编码蛋白序列由375 个氨基酸残基组成,原子总数为5 669 个,分子总质量为40 kDa,总分子式为C1770H2858N498O526S17。在组成蛋白的所有氨基酸中Leu 占比最高(16.5%),Tyr 含量最低(0.5%)。负电荷残基数大于正电荷残基数,为酸性蛋白。蛋白质具备弱亲水性,没有明显的疏水区段,所以可以推测CD14 蛋白没有跨膜区,并且已有研究结果表明mCD14 通过GPI 锚定结合在细胞膜上,符合跨膜区预测工具的分析结果[19]。详细内容见Fig.2。已知人体CD14 成熟蛋白有2 种存在形式:膜结合型和血浆游离型,详细见Tab.4 和Tab.5。
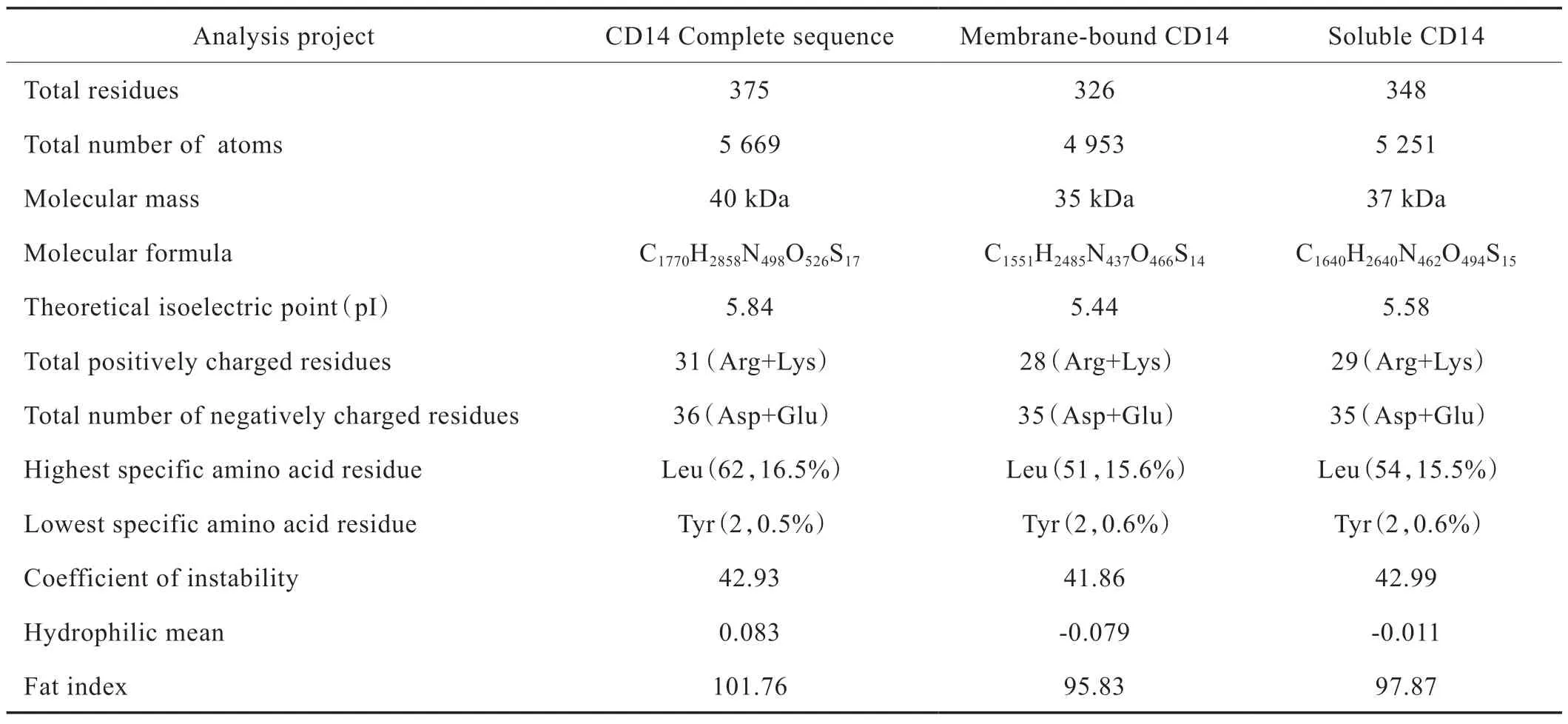
Tab.4 Analysis of physicochemical properties of human CD14 gene encoding protein
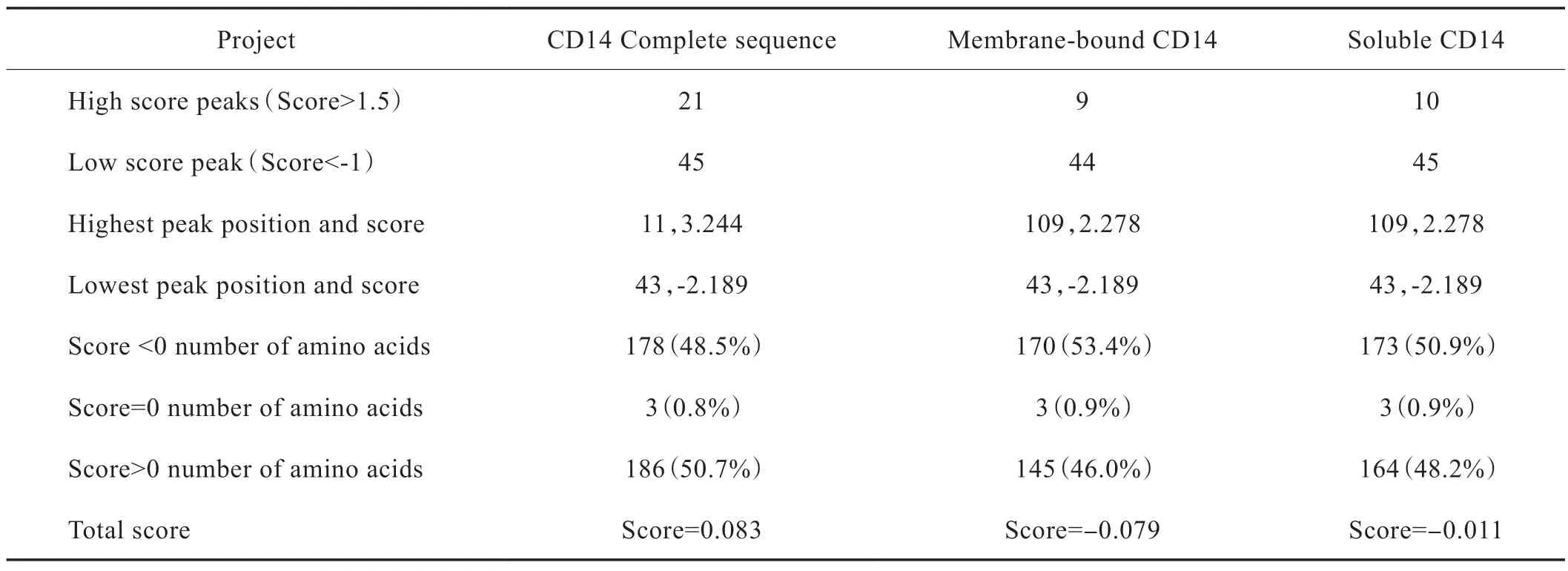
Tab.5 Hydrophilicity/ hydrophobicity analysis of human CD14 gene encoding protein
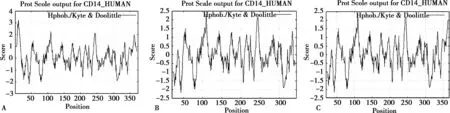
Fig.2 Hydrophilicity/ hydrophobicity analysis of protein encoded by CD14 gene(Kety and Doolittle algorithm)
2.2.2 人CD14 蛋白信号肽预测
基于神经网络和隐马可夫模型两种算法预测CD14 信号肽,结果一致,CD14 蛋白具有信号肽,为分泌蛋白,详细数据见Tab.6。
Tab.6 Human CD14 protein signal peptide prediction
2.2.3 CD14 蛋白亚细胞定位预测
采用亚细胞定位预测工具TargetP1.1 和WoLF PSORT,均表明人CD14 蛋白是种分泌蛋白,定位于细胞外基质(Tab.7)。

Tab.7 Prediction of human CD14 protein subcellular localization
2.2.4 人CD14 蛋白二级结构预测
采用SOPMA 在线预测工具对人CD14 蛋白质进行二级结构预测。SOPMA 预测结果表明人CD14 蛋白α螺旋(alpha helix)占整段蛋白序列的39.73%,β折叠(extended strand,β sheet)占比为12.27%(Fig.3)。
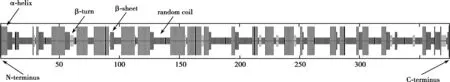
Fig.3 Human CD14 Secondary Structure Prediction(SOPMA)
2.2.5 同源建模法预测CD14 三级结构并进行模型质量评估
根据人CD14 蛋白序列在SWISS-MODEL 在线测序工具搜索预测模板,结果得到了PDB ID 为 4glp.1.A的预测模型,相似度为99.68%,GMQE 值为0.74,选用模板可靠性高。选用4glp.1.A 作为模板,而后用同源建模法得到Fig.4A。将得到的模型PDB 文件上传至SAVES v5.0 平台上进行在线评估。Fig.4B 是拉曼图来源于SWISS-MODEL,用于对模型的质量评估,评估结果显示95%以上的氨基酸残基都在合理的区域范围即Cα的二面角合理,表明该结构稳定。3D-1D 得分结果表明有88.39%的氨基酸残基3D-1D 得分大于等于0.2,高于80%氨基酸残基大于等于0.2 的标准,并且Fig.4C 显示低分部分较少,表明模型质量合格,三级结构预测结果稳定可靠。
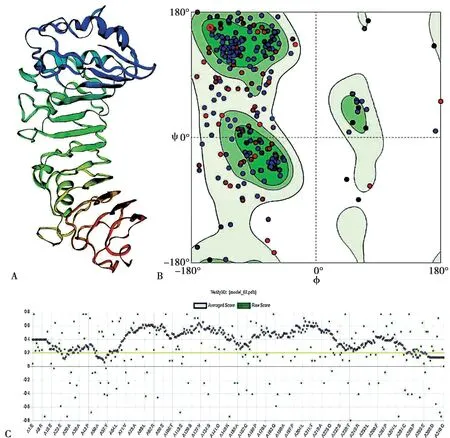
Fig.4 The use of homology modeling method to predict the tertiary structure of CD14 protein and its model quality evaluation results display
2.2.6 人CD14 互作网络及关联基因的功能注释
利用STRING 在线分析工具和Cytoscape 软件挖掘出以人CD14 基因编码蛋白为中心节点的蛋白质互作网络,并进行可视化,发现有10 个与CD14 互作的蛋白分别为:TLR2、LBP、LY96、ITGAM、ITGB2、MYD88、PLCG2、TLR1、TLR4、TLR6。其中CD14 蛋白与TLR4 基因关联最紧密(Fig.5A)。使用DAVID 数据库将CD14 与关联基因进行生物学功能注释,筛选最显著富集的生物学功能注释(P-value≤0.05 和Count≥4 genes)见Fig.5B,细胞对脂蛋白的反应(cellular response to lipoteichoic acid),细胞因子分泌的调 节(regulation of cytokine secretion),Toll 样受体信号通路(toll-like receptor signaling pathway),白细胞介素6 产生的正调控(positive regulation of interleukin-6 production),对脂多糖的反应(response to lipopolysaccharide),Toll 样受体4 信号通路(toll-like receptor 4 signaling pathway),脂多糖介导的信号通路(lipopolysaccharidemediated signaling pathway),肿瘤坏死因子产生的正调控(positive regulation of tumor necrosis factor production),MyD88 依赖的Toll 样受体信号转导途径(MyD88-dependent toll-like receptor signaling pathway),炎症反应(inflammatory response),先天免疫反应(innate immune response),质膜(plasma membrane),脂多糖结合(lipopolysaccharide binding),受体活性(receptor activity),其中CD14 在Toll 样受体4 信号通路和MyD88 依赖的Toll 样受体信号转导途径中发挥作用最强。
Fig.5 CD14 interacting protein and its functional annotation
3 讨论
单核苷酸多态性(single nucleotide polymorphism,SNP)是影响个体对疾病遗传易感的重要遗传因素[21],实验结果表明CD14-159C/T(rs2569190)等位基因TT的基因型与心血管疾病患者之间存在相关性,即TT 型患者心血管病发病率高于CT 型和CC 型。CD14 基因-159C>T(rs2569190)基因位点的改变可能影响心血管疾病的发生。
CD14 蛋白无跨膜区,完整CD14 序列有375 个氨基酸残基,mCD14 有326 个氨基酸残基,从第20位氨基酸残基开始以第345 位氨基酸残基结束,并且第345 位氨基酸残基为天冬酰胺,是糖基磷脂酰肌醇(glycosylphosphatidylinositol,GPI)的锚定结合位点,并且将CD14 通过GPI 锚定在单核细胞的核膜上,sCD14有348 个氨基酸残基,从第20 位至第367 位氨基酸残基,完整序列20 位氨基酸残基之前为蛋白质信号肽,367 位氨基酸残基之后的蛋白序列在蛋白成熟后被切除。mCD14 分子分布在巨噬细胞,树突细胞和中间粒细胞的表面,单核细胞可依据其表面CD14 和CD16 表达水平划分为不同亚群,并且Chang 等证实存在通过减少mCD14 分子水平而增加sCD14 分子水平的生物过程[22],这也为单核细胞不同亚群之间相互转换提供理论支持。其中CD14 在中间型单核细胞上高表达,并且中间型单核细胞含量增加常伴随有患心血管疾病的风险[23]。
CD14 是一种存在于单核细胞、巨噬细胞等细胞表面的白细胞分化抗原,由DAVID 富集通路结果可知,CD14 蛋白分子所参与的细胞组分是质膜,CD14 的分子功能是革兰阴性细菌细胞壁成分脂多糖(lipopolysaccharide,LPS)的受体,当LPS 与mCD14 结合时向单核细胞传递信号,刺激单核细胞分泌肿瘤坏死因子(tumor necrosis factor,TNF)、IL-1 等细胞因子,以及能够促进单核细胞对内皮细胞的黏附作用[24-25],而且异常的单核细胞的黏附和聚集很可能是急性心肌梗死和动脉硬化的发病原因[26],单核细胞分泌的炎症因子如TNF、IL-1 等可以正向调节CD14 基因,这将导致更高水平的CD14。DAVID 富集通路结果显示,与CD14 具有强关联性的生物学过程有Toll 样受体4 信号通路和MyD88 依赖的Toll 样受体信号转导途径,CD14 是LPS 的受体,其与 LPS 特异性结合后,可通过Toll 样受体4 以及MD2 蛋白向下游传递激活信号,从而启动单核-巨噬细胞系统(mononeuclear phagocyte system,MPS),并释放多种炎性细胞因子发生一系列反应[25],Kim 的小鼠TLR4 基因敲除实验同样得到了相同的结论,TLR4 作用增强与动脉壁炎症的发生具有强关联[27]。LPS 诱导单核细胞分泌炎症因子,阻遏CD14分子功能就可防止或中止发生LPS 性炎症反应等病理反应。并且CD14 在病原相关模式分子(pathogenassociated molecular patterns,PAMP)如细菌LPS 等刺激下被单核-巨噬细胞高表达,再激活体内单核细胞、内皮细胞和平滑肌细胞等靶细胞,使其释放炎症因子,引发炎症反应,正向调节CD14 的表达,并形成恶性循环,导致心血管疾病的发生[28-31]。
综上所述,基于生物信息学预测CD14 蛋白的理化性质和功能,CD14 基因表达水平升高可能在心血管疾病的发生发展过程中发挥重要作用;CD14 基因-159C/T(rs2569190)基因位点多态性与心血管疾病发生密切相关,其原因可能是TT 纯合子突变影响了CD14 基因的表达。因此,借助CD14 基因多态性和异常表达的筛查及早发现易患心血管疾病的人群,并尽早的进行预防,对于提升人民健康水平具有重大意义。
