四川卧龙国家级自然保护区雪豹地栖动物群落初探
2021-08-03周厚熊吴艳蓉王鹏彦施小刚李兆元
周厚熊 姜 楠 李 君 杨 虎 吴艳蓉 王鹏彦 施小刚 李兆元*
(1.西南林业大学生物多样性保护学院,昆明,650224;2.昆明逸境生态工程咨询有限公司,昆明,650224;3.四川卧龙国家级自然保护区管理局,阿坝,623006)
雪豹(Pantherauncia)隶属于食肉目(Carnivora),猫科(Felidae),豹属[1],全球种群数量7 463—7 980 只[2],国外分布于阿富汗、不丹、印度、哈萨克斯坦、吉尔吉斯斯坦、蒙古、尼泊尔、巴基斯坦、俄罗斯、塔吉克斯坦和乌兹别克斯坦[3],国内分布于西藏、新疆、青海、甘肃、四川、云南[4],其中四川卧龙国家级自然保护区是雪豹全球分布的东南边界[5]。作为山地生态系统的旗舰种和顶级捕食者,雪豹在维持生态系统的健康和稳定中起着极其重要的作用[6]。到目前为止,雪豹所生活的地栖动物群落构成及其在群落中的生态学作用还不清楚。
保护生物学理论认为,顶级捕食者的存在是判断一个生态系统健康与否的重要标志[7]。然而,顶级捕食者如何影响生态系统健康需要从群落角度去认识。Odum[8]、Ricklefs[9]以及Putman[10]认为,群落是物种通过生态学关系(如:捕食、竞争、共生等)结成的整体。在这个整体中,生态学关系支撑物质和能量在物种间的传递,以实现生态系统的功能。因此,群落不是一群物种的简单集合;它是物质和能量传递过程中涉及的物种的集合。生态学关系是理解群落结构的关键。然而,传统的生态学研究难以采集物质和能量作为通货,同时量化各个物种间的生态学关系[11]。由于所有种间生态学关系都需通过物种种群有规律地出现在特定地理空间中才得以实现,因此物种的生态学关系必然呈现在物种的空间关联之中。为此,本研究利用四川卧龙国家级自然保护区红外相机记录,通过种间空间关联构建雪豹所生活的地栖动物群落,从群落角度分析该物种的生态学作用,探讨它对维持高山生态系统的意义,从而为保护管理工作提供科学建议。
1 研究区域概况
卧龙国家级自然保护区(30°45′—31°25′N,102°52′—103°25′E)位于四川省汶川县,属成都平原向青藏高原过渡的高山深谷地带,面积约2 000 km2(图1)。保护区地势东南低、西北高,海拔1 150—6 250 m。保护区内有耿达河、皮条河、正河、西河和中河,均属岷江水系。地表景观随海拔上升依次为常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、针叶林、亚高山灌丛和草甸,以及高山流石滩[12]。保护区内分布有大熊猫(Ailuropodamelanoleuca)、雪豹、小熊猫(Ailurusfulgens)、黑熊(Ursusthibetanus)、黄喉貂(Martesflavigula)、藏酋猴(Macacathibetana)、川金丝猴(Rhinopithecusroxellana)、羚牛(Budorcastaxicolor)、岩羊(Pseudoisnayaur)、林麝(Moschusberezovskii)、绿尾虹雉(Lophophoruslhuysii)、红喉雉鹑(Tetraophasisobscurus)、血雉(Ithaginiscruentus)、白马鸡(Crossoptiloncrossoptilon)、藏雪鸡(Tetraogallustibetanus)以及金雕(Aquilachrysaetos)等珍稀物种。
2 研究方法
2.1 数据采集
2017年4月—2018年4月,在保护区内皮条河东岸随机选择3个20 km2的样区(图1),所选样区海拔1 749—4 430 m。样区涵盖所有生境类型,包括常绿阔叶林、落叶阔叶林、针阔混交林、针叶林、灌丛、高山草甸、流石滩。用ArcGIS 11软件将样区划分成60个1 km×1 km的公里网格,每个网格为1个采样单元。将网格内占地>50%的生境类型定义为采样单元的生境类型,并在其中安装1台红外相机(易安卫士L710)。相机固定在稳固的树干或岩石上,距地面30—50 cm,并用枯枝落叶进行隐蔽。同时,记录位点的经纬度、海拔、坡位、坡度、坡向、地物类型、森林演替状态、植被高度和植被盖度。采用Wei等[13]采集生境因子的方法。每台相机配置16 GB内存卡,相机时间设定为北京时间,灵敏度设定为中,拍摄间隔时间1 s,触发后连续拍摄3 次,随后录像20 s。电池和内存卡每隔3个月更换1次。物种鉴定参考《中国鸟类野外手册》[14]和《四川兽类原色图鉴》[15]。在安装相机以及更换相机电池及内存卡时,数据采集者对相机位点周围进行勘察,记录所发现的动物实体及痕迹。痕迹鉴定参考《中国兽类踪迹指南》[16]。所有照片数据导入电脑,经整理后录入Microsoft Excel,以供分类、计算和汇总。
2.2 物种相关性分析
对各位点照片进行物种鉴定,获取物种存在与否的二分型数据。针对这种二分型数据,采用phi系数对种间关联度进行衡量[17]。
在进行Phi系数计算时,首先建立2×2列联表(表1)。将全部相机位点分为4组:第1组为物种X和物种Y均出现的相机位点,记为A组;第2组是物种X不出现,但物种Y出现的位点,记为B组;第3组是物种X出现,物种Y不出现,记为C组;第4组是物种X和物种Y均不出现的位点,记为D组。计算各组位点数,填入列联表。表1中,N=A+B+C+D,即:总相机位点数。

表1 Phi系数2×2列联表
Phi系数(rφ)计算公式如下:
(1)
rφ取值0—1:接近0,表示种间空间关联性弱;接近1,表示关联性强。如果(AD-BC)>0,表明种间存在正关联;如果(AD-BC)<0,则种间存在负关联。在本研究中,2个物种共同出现于相同空间频率越高,phi系数越大。在捕食者-猎物关系中,Phi系数越大,表明捕食者物种对猎物物种的捕食压力越大以及猎物物种对捕食者物种的生存意义越大;在对相同资源竞争的物种中,则表明种间的竞争越激烈。
进一步用卡方检验求取上述phi系数值的显著性,公式如下:
(2)
当χ2≥3.84时,P≤0.05,表明计算所获rφ值有意义,是生态学关系导致的空间关联,可用于研究分析。当χ2<3.84时,P>0.05,表明计算所获rφ值无意义,关联现象是随机因素导致的,不能用于研究分析。
用lambda系数帮助判断存在直接空间关联的食肉目动物间的实际生态学关系,计算过程参照李兆元等[17]。
2.3 群落构建
依据上述种间关联性计算,找出具有显著正关联的种对。通过种对关系,将物种构建成空间关联网络。最后,通过生态学文献及野外工作中发现的证据,将种对空间关联关系还原成生态学关系,从而获得群落物种网络。
2.4 种间关联多样性
群落的总体稳定性和抗干扰力与群落的整体复杂性紧密相关。本研究用香农-威纳指数(Shannon-Wiener)来衡量群落的整体复杂性,计算如下:
(3)
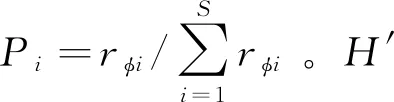
2.5 生态因子分析
采用卡方吻合度检验分析雪豹分布的生态制约因子。分析群落分布的生态制约因子涉及多个物种,各生态因子也涉及多种表达,如:生境类型。因此,采用克莱姆系数(C)进行检验,计算过程参照李兆元等[17]。
3 结果与分析
3.1 地栖动物区系
在60个公里网格中,红外相机的总工作时间为16 966台日,获得清晰照片7 120张,鉴定出6目15科35种地栖动物。其中,兽类(Mammalia)有5目14科26种,分别为喜马拉雅旱獭(Marmotahimalayana)、豪猪(Hystrixbrachyura)、藏鼠兔(Ochotonathibetana)、藏酋猴、川金丝猴、狼(Canislupus)、赤狐(Vulpesvulpes)、黑熊、大熊猫、小熊猫、黄喉貂、石貂(Martesfoina)、黄鼬(Mustelasibirica)、香鼬(M.altaica)、猪獾(Arctonyxcollaris)、花面狸(Pagumalarvata)、豹猫(Prionailurusbengalensis)、雪豹、野猪(Susscrofa)、林麝、毛冠鹿(Elaphoduscephalophus)、水鹿(Rusaunicolor)、羚牛、中华鬣羚(Capricornismilneedwardsii)、中华斑羚(Naemorhedusgriseus)和岩羊;鸟类(Aves)1目1科9种,分别为雪鹑(Lerwalerwa)、红喉雉鹑、藏雪鸡、血雉、红腹角雉(Tragopantemminckii)、勺鸡(Pucrasiamacrolopha)、绿尾虹雉、白马鸡和红腹锦鸡(Chrysolophuspictus)。此外,还记录到山羊、黄牛、牦牛和村民。在35种野生动物中,食肉目物种丰度最高,含6科13种;其余依次为偶蹄目(Artiodactyla)4科8种,鸡形目(Galliformes)1科9种,啮齿目(Rodentia)2科2种,灵长目(Primates)1科2种,兔形目(Lagomorpha)1科1种。
雪豹在8个相机位点中出现,海拔范围4 072—4 430 m,平均海拔4 252 m;属于高山苔原生物相,生境景观类型包括灌丛、流石滩和高山草甸。海拔分布重叠或部分重叠的物种有18个,包括喜马拉雅旱獭(海拔范围:3 718—4 430 m,平均4 125 m)、赤狐(3 292—4 430 m,3 980 m)、香鼬(4 303—4 338 m,4 320 m)、石貂(4 303—4 338 m,4 249 m)、豹猫(1 979—4 303 m,3 004 m)、猪獾(2 473—4 430 m,3 394 m)、岩羊(2 473—4 430 m,4 024 m)、藏鼠兔(2 473—4 430 m,4 151 m)、斑羚(1 749—4 430 m,2 924 m)、血雉(3 042—4 303 m,3 733 m;灌丛、流石滩、草甸)、雪鹑(3 186—4 430 m,4 048 m;灌丛、流石滩、草甸)、红喉雉鹑(3 718—4 052 m,3 912 m;灌丛)、藏雪鸡(4 152—4 300 m,4 226 m)、绿尾虹雉(3 093—4 338 m,3 841 m;灌丛、流石滩、草甸)。
卡方吻合度检验结果显示,雪豹分布与海拔、坡向、地物类型和植物高度存在显著相关性(表2),分布于海拔4 000 m以上、西向、灌丛和流石滩区域,回避低海拔高植被区域。植被盖度、坡度和坡位对该物种的分布没有显著性影响。
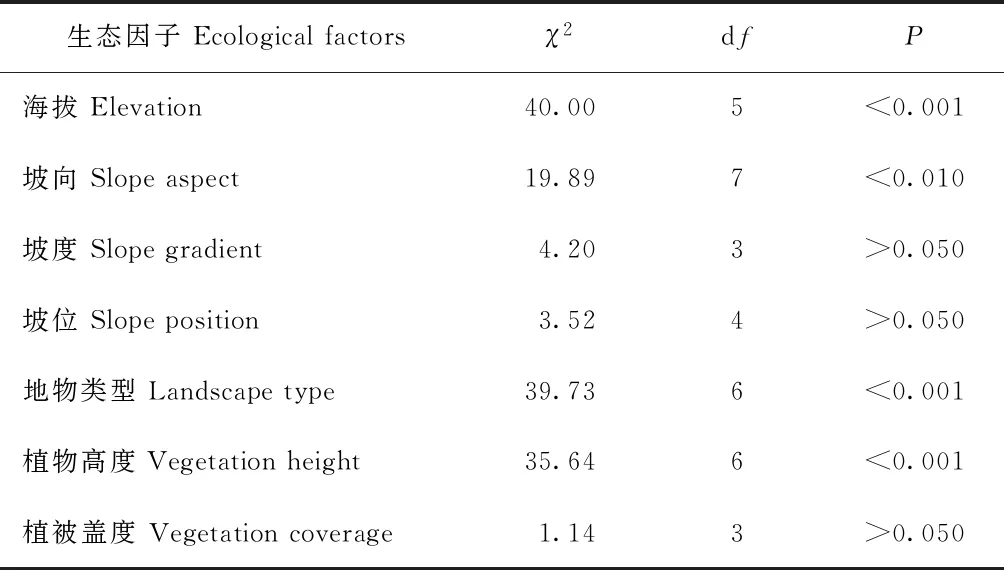
表2 雪豹分布与各生态因子相关性检验结果(卡方吻合度检验)
3.2 地栖动物空间关联网络
3.2.1 关联网络构成
Phi系数计算结果显示,35种同域分布的地栖动物中,有13种与雪豹存在直接或间接空间正关联,包括喜马拉雅旱獭、赤狐、香鼬、石貂、岩羊、藏鼠兔、血雉、白马鸡、猪獾、雪鹑、红喉雉鹑、绿尾虹雉和藏雪鸡。这些物种与雪豹在海拔分布上重叠,共同构成空间关联物种网络(图2)。网络中共有14个物种,占区系物种数40%。这些物种结成38个种对,关联多样性(香农-威纳指数)为7.30。在这个网络中,赤狐与血雉的关联度最强(0.67),其次是红喉雉鹑和白马鸡(0.64)、雪豹和岩羊(0.63);与雪豹关联度最高的是岩羊(0.63),其次是雪鹑(0.59)和石貂(0.59)。网络结构大致分为2层:外层是藏雪鸡、香鼬、藏鼠兔、白马鸡、红喉雉鹑、猪獾和雪鹑,每个物种的种对最多4个,关联系数平均为0.47;内层为绿尾虹雉、喜马拉雅旱獭、血雉、赤狐、石貂、岩羊和雪豹,各物种的种对数至少7个,平均关联系数是0.48。内、外层关联系数无显著性差异(F=0.75,df=55,P>0.05;t=0.98,df=55,P>0.05),但种对数存在显著性差异(χ2=4.57,df=1,P<0.05),表明内层物种对维持网络稳定有主要作用。其中,雪豹为大型食肉类,在本网络中是最大的捕食者,可能是顶级捕食者;赤狐为中型捕食者。这2种捕食者没有发生直接空间关联,而是通过共同的关联物种(包括绿尾虹雉、喜马拉雅旱獭、石貂和岩羊)发生间接关联,可能表明由于食性竞争而相互回避。雪豹与石貂、喜马拉雅旱獭、岩羊、雪鹑空间关联强(rφ>0.5),表明雪豹对这些物种可能依赖性强。赤狐与白马鸡、红喉雉鹑、血雉、岩羊,以及喜马拉雅旱獭也存在很强的空间关联,可能对这些物种存在强的依赖性。值得注意的是,2种中大型食草动物岩羊、喜马拉雅旱獭同时与雪豹、赤狐产生显著空间正关联,对支撑雪豹和赤狐的生存均表现出重要意义;小型食肉动物石貂也同时与雪豹、赤狐产生显著空间关联,表明食肉动物之间可能存在相互捕食或一方为另一方创造有利条件的关系。
3.2.2 种间非对称关联
上述物种网络中,体形大小迥异但存在显著空间正关联的捕食物种种对有雪豹-石貂、雪豹-香鼬、赤狐-石貂、赤狐-猪獾、石貂-香鼬。Lambda系数检验结果显示:石貂能预测雪豹出现(λB>0.01,P≤0.05),表明雪豹捕食石貂;香鼬能预测石貂出现(λB>0.03,P≤0.05),表明石貂捕食香鼬;猪獾能预测赤狐出现(λB>0.03,P≤0.05),表明赤狐捕食猪獾;雪豹与香鼬、赤狐与石貂不存在空间非对称关联(λB<0,P>0.05)。
3.2.3 物种网络与生态因子的关系
网络中的物种总体随海拔升高出现频次增加,其中,在3 531—4 430 m区段内出现的频率占总频率81.30%,为主要分布区段。克莱姆系数检验结果(表3)显示,该网络的分布与坡向、生境类型、森林演替状态、植被高度和植被覆盖度具有显著相关性,坡度和坡位没有显著相关性;网络中的物种在灌丛、草甸(或草地)、流石滩以及无林空间中出现的频率最高,占78.05%;表明这些生境类型是网络赖以存在的基础;这些生境类型主要分布于海拔3 718—4 430 m区段中。
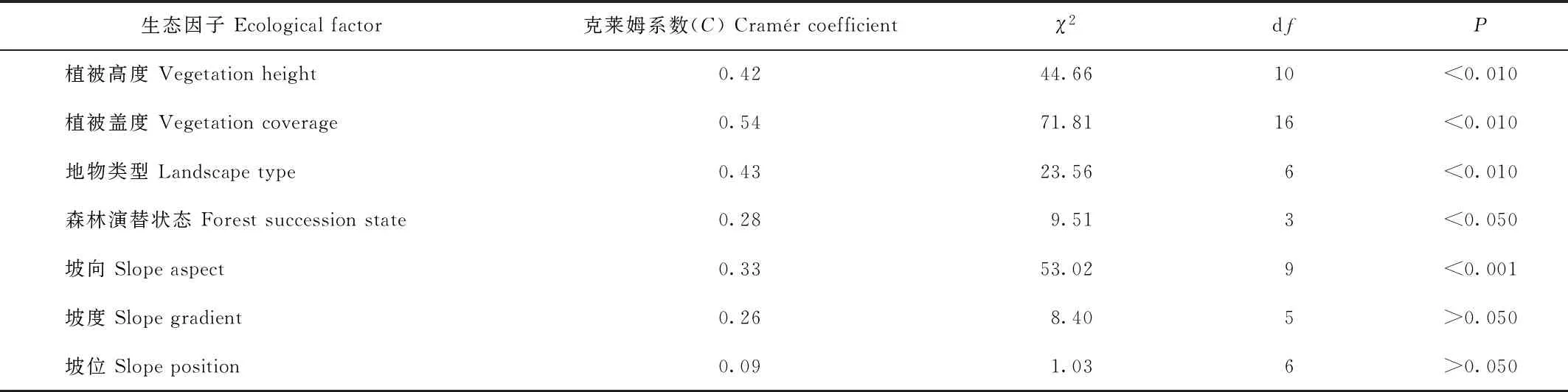
表3 物种分布网络与生态因子的相关关系
综上所述,该物种网络分布于海拔3 718 m以上,植被稀疏的灌丛、草甸(或草地)和流石滩生境中。网络由高海拔物种(旱獭、雪豹、香鼬、石貂、白马鸡、红喉雉鹑、藏雪鸡)以及海拔广布种的高海拔种群(赤狐、岩羊、藏鼠兔、血雉、猪獾、雪鹑、绿尾虹雉)构成。
4 结论与讨论
4.1 数据采集方法及质量评估
以往关于物种空间关联的研究主要采用样线法收集动物在沿线留下的痕迹(如毛发、粪便、取食痕迹等),从而获取动物在特定空间中出现与否的数据[18-19]。这种方法存在以下问题:①考虑到数据可比性,所有地理空间中的数据需要同时采集。但是,样线法无法使研究者同时进行大面积数据采集。②如果动物未留下痕迹,所获数据可能非常不完整。③在确定不同地点发现的痕迹是否属于分布于同一地理空间时,样线法无法提供客观的判断依据。随着红外相机技术的出现,这些问题逐步得到有效解决。第一,红外相机可以长时间连续工作,准确提供相机位点附近物种的出现情况[20-21],同时布设多台相机,可以有效解决上述第1个问题。第二,红外相机在无人操作的情况下可以自动触发拍照,获得动物活体照片,很大程度解决上述第2个问题。第三,相机位点可以作为客观参照物,位点附近出现的动物痕迹以及相机拍摄到的动物可以视为相同地理空间中的存在物,这样解决了上述第3个问题。
与其他研究方法一样,红外相机技术也存在特有问题。首先,红外相机依靠温血动物身体温度与环境温度的温差来触发。冷血动物(两栖类Amphibia、爬行类Reptilia)身体温度与环境温度相似,无法触发相机拍照,因此该技术无法获取冷血动物的照片记录。其次,李晟等[20]研究发现红外相机对小型哺乳动物图像捕获率偏低。这种技术缺陷可能会影响研究结果。例如,魏万红等[22]研究表明香鼬主要猎物是藏鼠兔;但本研究结果显示,这2个种没有显著空间关联性。藏鼠兔是小型哺乳动物,可能存在红外相机对其影像捕获率偏低的问题,但有待验证。本研究所获数据中,小型哺乳动物似乎存在整体上的代表性偏低问题。空间关联计算中,2个物种共同出现在相同地理空间中的频率是建立种间关联的基础,受物种种群密度影响。因此,2项研究结果的差异可能由红外相机的技术缺陷导致。当然,也可能是数据采集期间藏鼠兔实际种群密度偏低所致。然而,这个缺陷没有影响中大型食肉动物雪豹的种间关系分析。
4.2 统计学方法选择
地栖动物基本都有家域[23]。部分物种为独居生活类型,其他为群居类型。如果相机布设在某一个体或群体的家域核心区,相同个体在相机探测区域反复活动,活动频率与种群密度的换算关系因动物生活类型不同而异。现有红外相机技术无法进行图像识别,研究者凭肉眼对个体进行识别所获结果不具备可靠性,尤其是对夜行性动物的识别(图像清晰度低,黑白色)。因此,特定位点相机所获“独立有效照片数”[20]不能用作物种密度的指示,而只能代表物种出现与否。针对这种二分型数据,只能选择非参数统计中的Phi系数来量化2个物种的空间关系[17]。
在一个群落中,食肉目物种常常不止一种。当体形差异较大的2种食肉目动物出现空间关联时,关联性可能是它们在相同地理空间中共同取食某种猎物(对共同食物的需求)所致,也可能是体形大的物种捕食体形小的物种(捕食关系)所致。此时,在没有相关文献支持下,难以判断它们的种间生态学关系。然而,在捕食关系中,被捕食者倾向于在其生境内随机活动,以摆脱被捕食;捕食者则依据被捕食者的活动来决定自己的活动空间,以便获得捕食机会。因此,两者的空间关联性不对称,可以用Lambda系数检验预测2个物种的空间出现[17]。
4.3 地栖群落构成
本研究所获数据均为地栖动物空间分布数据。基于这些数据建立的物种网络是个地栖物种网络,通过生态学关系关联起来形成一个地栖动物群落。雪豹是地栖动物,它的所有生态学关系发生在这个地栖动物群落中。
Schaller等[24]、刘楚光等[25]通过粪便(样本量分别为206、644 份)研究发现,喜马拉雅旱獭、岩羊是雪豹的重要食物来源,2种猎物的残留物在粪便中出现频次相当,说明喜马拉雅旱獭和岩羊在雪豹食谱中的重要性相似。本研究结果显示,雪豹与喜马拉雅旱獭以及雪豹与岩羊的空间关联系数相似,分别为0.57和0.63;表明雪豹对喜马拉雅旱獭和岩羊的捕食压力比较接近,与上述研究结果一致。Oli等[26]在尼泊尔的研究发现,雪豹捕食小型食肉类石貂、伶鼬(Mustelanivalis),在213份雪豹粪便中,石貂的残留物有8份,伶鼬的残留物有10份,说明雪豹作为机会主义捕食者也捕食中小型食肉类。本研究结果显示,石貂能预测雪豹出现(λB>0.01,P≤0.05),表明雪豹捕食石貂,与上述结果一致。香鼬不能预测雪豹出现,表明雪豹不会规律性地捕食香鼬。从雪豹与喜马拉雅旱獭的捕食关系和香鼬与喜马拉雅旱獭较强的空间关联性来推测,雪豹与香鼬之间可能是掠食关系,即雪豹捕食喜马拉雅旱獭后,香鼬再利用剩余部分。Oli等[26]研究发现,雪豹捕食鸡形目鸟类,但鉴于当年的技术条件无法鉴定是哪一种。根据研究地鸡形目鸟类的分布,推测其可能是藏雪鸡、暗腹雪鸡(Tetraogallushimalayensis)或石鸡(Alectorischukar)。本研究结果表明,雪豹与藏雪鸡空间关联性(0.47)强,捕食关系可能促使它们共同出现在相同空间的概率增加。雪豹与雪鹑、绿尾虹雉之间的空间关联可能由类似的原因导致。基于以上分析,获得雪豹与各直接关联物种的种间生态学关系,如图2所示。
4.4 群落抗干扰能力
图2群落中,食草动物有岩羊、喜马拉雅旱獭和藏鼠兔,杂食动物有绿尾虹雉、红喉雉鹑、白马鸡、雪鹑、血雉、藏雪鸡和猪獾,食肉动物有雪豹、赤狐、石貂和香鼬。每个营养级上都有2个以上的物种。当同一营养级中一个物种出现种群密度下降时,其他物种可以替补其生态学功能,由此为群落提供应对外来冲击的韧性。如:当雪豹因环境动荡出现种群密度下降时,同样处于捕食者的赤狐可以替代雪豹控制食草动物数量;反之亦然。通过这种机制,食草动物种群数量可以受到持续调控,避免草场被过度利用,群落得以有效维持。
4.5 雪豹在群落中的角色
雪豹是群落的顶级捕食者,被保护生物学家视为高海拔旗舰物种[27]。本研究结果(图2)表明,雪豹和赤狐是群落中重要捕食物种,像2只手控制着群落中的其他物种。雪豹捕食藏马鸡、香鼬、石貂、喜马拉雅旱獭、绿尾虹雉、岩羊和雪鹑,共7种,占群落物种数50%,其中,石貂(0.59)、喜马拉雅旱獭(0.57)、岩羊(0.63)和雪鹑(0.59)为主要食物物种。赤狐捕食白马鸡、红喉雉鹑、猪獾、血雉、绿尾虹雉、岩羊、石貂和喜马拉雅旱獭,共8种,占群落物种数57%;从相关系数看,白马鸡(0.53)、红喉雉鹑(0.60)、血雉(0.67)、岩羊(0.55)和喜马拉雅旱獭(0.54)为主要食物物种。二者共同的食物物种有:岩羊、喜马拉雅旱獭、石貂、绿尾虹雉。从猎物物种总数来看,赤狐比雪豹多1种。但值得注意的是,从体型上看,赤狐体重5—7 kg不太可能捕食成年岩羊(60—75 kg)[15],在群落中仅有雪豹能对成体岩羊起到控制作用。因此,作为顶级捕食者雪豹在高海拔群落中具有不可替代的作用,应该加强保护与研究。
综上所述,本研究认为卧龙国家级自然保护区雪豹地栖动物群落作为一个实体客观存在,群落中各营养级都有2个以上的物种,同级物种的生态角色可以相互替换,表明卧龙雪豹群落是一个健康的群落,具备继续演化的能力。
致谢:感谢四川省林业和草原局古晓东的大力支持;卧龙国家级自然保护区胡强、王茂麟、林红强的倾力协助;剑桥大学李梦烨女士帮助修改英文摘要!
