水分胁迫/复水对谷子光合特性及产量影响
2021-07-29任传友姜卓群苏小琁米前川高西宁
任传友 姜卓群 苏小琁 米前川 王 婧 李 玥 高西宁
(沈阳农业大学农学院, 沈阳 110866)
引 言
干旱是水分收支长期不平衡导致的持续水分短缺现象[1],对作物的生长发育及产量形成具有重要影响[2-4]。谷子是我国主要的大田经济粮食作物之一,在我国北方干旱地区分布广泛,曾是我国北方干旱地区的主要作物[5]。谷子具有良好的耐旱性,是应对未来日趋严重的水资源短缺的战略储备作物,也是当前我国旱区农业种植结构优化调整的首选作物[6]。随着全球气候变化,干旱风险增大[7-8],我国东北和华北干旱化趋势显著[9],对谷子的高产稳产带来严峻挑战。明确谷子对干旱的响应规律,对谷子田间科学管理具有重要指导作用。
光合作用是作物生长发育和产量形成的基础和保证。光合过程对水分胁迫的响应与干旱强度、持续时间、发生时间及作物发育阶段有关[10-12]。轻度干旱条件下,作物可能不受影响,甚至可能改善作物生长、提高产量和水分利用效率[13-14],利用该特性可通过亏缺灌溉和节水农业措施提高可持续发展[15-18],但在严重干旱条件下,作物表现为生长限制或受到伤害[16]。复水后,遭遇水分胁迫的作物通常表现为生长加速和光合能力提高,这在一定程度可补偿干旱的影响[19-22]。补偿效应高低与作物发育阶段、水分胁迫强度及持续时间有关[20-21,23],如冬小麦花前短期轻旱能在不显著降低产量的情况下提高水分利用效率,在抽穗期遭受干旱复水后补偿效应更高[24],灌浆期轻度水分胁迫可实现节水和高产统一[25]。农谚常说,谷子最怕“胎里旱”,谷子在穗分化期遇到干旱,会影响穗枝梗和小穗分化,形成穗小、秃尖等现象,穗粒数减少。但也有研究表明:在谷子的各发育阶段,干旱胁迫对灌浆期的影响最明显[26]。谷子在生长过程中经常会经历不同季节、不同发育阶段的干湿变化,其抗旱性不仅体现在对干旱状态的耐受程度上,还应包括胁迫状态解除后的恢复能力。因此,谷子干旱后复水机理研究对其抗旱性改良具有重要意义。光合作用的改变是水分胁迫的直接体现,并最终与产量形成相关联。
本研究利用大田控水试验研究水分胁迫/复水对谷子孕穗开花期和灌浆期光合特性的影响,分析其在产量形成中的作用,期望为干旱条件下谷子亏缺灌溉及农业节水措施的制定提供理论依据。
1 材料与方法
1.1 试验材料及栽培措施
试验于2020年在沈阳农业大学北山科研基地遮雨棚内进行。地块平坦,土质为棕壤,地力中等,土壤容重为1.25 g·cm-3,重量田间持水量为30%,选用辽宁省广泛种植的谷子品种大金苗,种子来源于辽宁省建平县农业技术推广中心。条播种植,垄宽0.6 m,留苗密度为45 万株·hm-2。当地常规施肥水平,采用28-16-11复合肥(55%)配施硫酸钾肥(含50% K2O),施肥水平是纯氮为98 kg·hm-2,磷肥P2O5为56 kg·hm-2,钾肥K2O为140 kg·hm-2。5月5日播种,正常田间管理,9月21日收获。
1.2 试验设计
试验共设置15个小区(12个水分胁迫处理小区,3个对照(control check,CK)小区,每个小区6垄,垄长为2 m,垄宽为0.6 m,各小区间采用防水聚氯乙烯板隔断,埋深为0.6 m。考虑到谷子不同发育阶段的需水差异,孕穗开花期(B)比灌浆期(F)需水更多,孕穗开花期设置45%(H,高度水分胁迫)和55%(L,轻度水分胁迫)田间持水量的水分胁迫处理,灌浆期高度、轻度水分胁迫水平分别为40%和50%田间持水量,持续时间分别为7 d,14 d和21 d(分别标记为07,14,21),胁迫处理后复水(R),灌水达田间持水量的65%(孕穗开花期)和60%(灌浆期)。CK小区土壤水分含量在孕穗开花期恒定在田间持水量的65%,该阶段前后分别为55%和60%,水分胁迫处理小区在其他时段保持与CK小区的土壤水分含量相同。为尽可能保持土壤水分恒定,遵循少灌多次原则,2 d灌水1次,按20 cm 深度测算达到目标土壤水分需灌溉的水量。胁迫处理结束当日和复水7 d后的上午采用LI-6400测定倒二叶的光响应曲线,3个重复。每个小区收获3垄,每垄单独测产,取平均值为该小区产量。
光响应曲线在CO2浓度为400 μmol·mol-1下测定,共设置15个光量子通量密度(FP)水平,分别为2000,1500,1200,1000,800,600,400,200,100,80,60,40,20,10 μmol·m-2·s-1和0。采用发育阶段(B或F)胁迫强度(H或L)-持续时间(07,14或21)[-R]方式标记测定的光合数据,如BH-14表示孕穗开花期45%田间持水量控水14 d后测定的数据,FL-07-R表示灌浆期50%田间持水量控水7 d、复水7 d后测定的数据。
1.3 计算指标
植物的光合作用过程受气孔因素和非气孔因素的共同影响。气孔因素的主要表征是气孔开度大小,影响CO2能否顺畅地进入气孔内部,从而决定参与光合作用过程的CO2含量及浓度。非气孔因素表现为参与光合作用的酶含量和活性,决定进入气孔内部的CO2能否被顺利转化为光合产物。通常植物在受到胁迫条件下,气孔导度会降低,酶含量和活性下降,光合过程受阻。判断气孔限制值的两个因素是Ci和Ls[27],其中气孔限制值Ls表示为
Ls=1-Ci/Ca。
(1)
式(1)中,Ci为胞间CO2浓度(单位:μmol·mol-1),Ca为大气CO2浓度(单位:μmol·mol-1)。
水分利用效率是评价水分亏缺条件下植物生长适宜度的综合指标之一,反映植物耗水与干物质生产之间的关系。本研究的叶片水平上水分利用效率采用定义
EW=RP/RT。
(2)
式(2)中,EW为水分利用效率(单位:μmol·mmol-1),RP为光合速率(单位:μmol·m-2·s-1),RT为蒸腾速率(单位:mmol·m-2·s-1)。
1.4 数据处理
采用t检验法进行处理间产量差异分析并采用如下规则标记:①将产量数据平均值从大到小依次排序;②首先在最大的数值上标记字母“a”;③依次序将后面的数值与最大的数值进行比较,如差异不显著,仍标记字母“a”,否则标记字母“b”;④以第1个标记字母“b”的数值作为新的最大值,依次序将后面的数值与之进行比较,如差异不显著,仍标记字母“b”,否则标记字母“c”;依次类推,直至所有的数值标记完毕。显著性水平设定为p=0.05。
2 结果分析
2.1 不同发育阶段水分胁迫/复水对产量影响
不同发育阶段水分胁迫/复水对产量的影响与胁迫强度及持续时间有关,同时发育阶段间存在差异(图1)。谷子在孕穗开花期遭遇持续7 d和14 d的轻度水分胁迫,与CK相比产量有所下降,但差异并不显著,随着持续时间延长到21 d,产量显著降低。随胁迫增强,产量影响加剧,高度水分强迫持续14 d可造成产量显著降低,进一步延长到21 d,可导致产量下降32.4%。在相同处理条件下,灌浆期水分胁迫/复水对产量的影响比孕穗开花期更明显,灌浆期轻度水分胁迫持续14 d可造成产量降低22.1%,FH-21处理组产量仅为CK的47.1%。
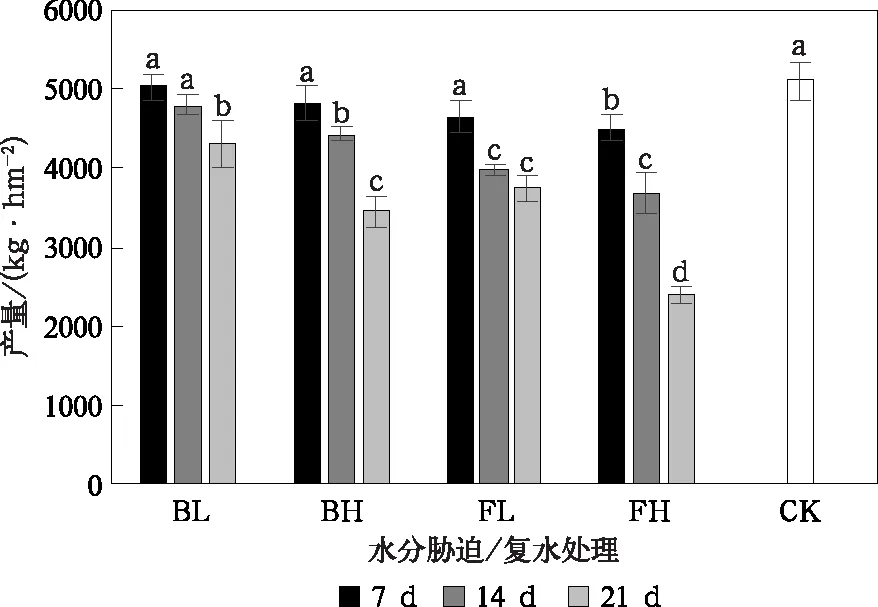
图1 各水分胁迫/复水处理的产量 (短线表示±1倍标准差)Fig.1 Effect of water stress/rewatering on foxtail millet yield (short line denotes ±1 time standard deviation)
2.2 水分胁迫/复水对孕穗开花期叶片光合速率和水分利用效率影响
水分胁迫对光合速率和水分利用效率的影响与胁迫强度和持续时间有关(图2和图3)。在孕穗开花期,轻度、高度水分胁迫处理7 d均可显著降低中高光强下叶片光合速率和提高水分利用效率,且随着胁迫增强光合速率降幅增大(图2),但水分利用效率增幅变小(图3)。胁迫时间延长至14 d时,BL-14光合速率虽仍低于CK,但二者差异已不明显,而BH-14仍显著地低于CK(图2);对于水分利用效率,BL-14高于CK,BH-14低于CK(图3)。当水分胁迫进一步延长到21 d时,轻度、高度水分胁迫对光合速率和水分利用效率的影响无明显差异,均低于CK。
复水后光合速率和水分利用效率恢复能力亦与水分胁迫强度和持续时间有关。持续7 d的水分胁迫处理组在复水7 d后,中高光强下的光合速率和水分利用效率显著高于CK,且BH-07-R比BL-07-R恢复能力更强(图2和图3)。对于14 d的处理组,复水7 d后光合速率亦有所恢复,高度水分胁迫处理组恢复更明显,但二者光合速率均未超过CK(图2),水分利用效率基本与CK持平(图3)。持续21 d处理组在复水后,光合能力恢复明显受抑制,复水7 d后二者的光合速率显著地低于CK,且水分胁迫越强,恢复能力越弱(图2),但BH-21-R在中低光强下水分利用效率显著提高(图3)。

图2 孕穗开花期光合速率对光量子通量密度的响应Fig.2 Responses of photosynthetic rate to photosynthesis photon flux density at booting and flowering stage
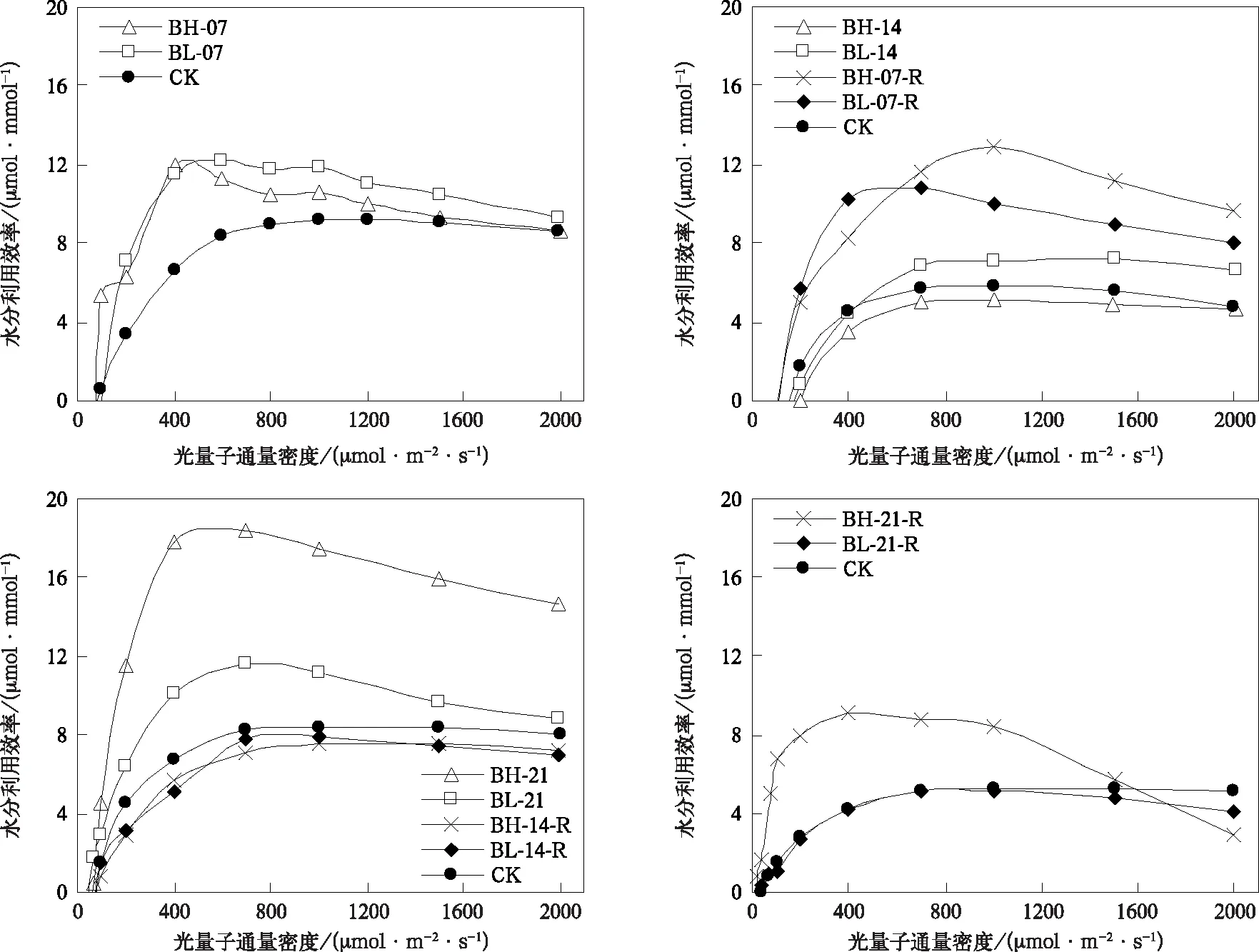
图3 孕穗开花期水分利用效率对光量子通量密度的响应Fig.3 Responses of water use efficiency to photosynthesis photon flux density at booting and flowering stage
气孔行为是制约光合作用强度的重要因素之一[28],图4是孕穗开花期不同轻度、高度水分胁迫及其复水后处理后叶片气孔导度对光量子通量密度的响应,总体上孕穗开花期水分胁迫可导致叶片气孔导度gsw(单位:mol·m-2·s-1)下降。由图4可知,与CK相比,轻度、高度水分胁迫7 d可导致光量子通量密度高于1000 μmol·m-2·s-1时叶片gsw下降49.6%~50.7%和66.8%~69.8%;当胁迫延长到14 d,二者的降幅分别为17.8%~24.4%和35.6%~42.5%;随着胁迫时间进一步延长到21 d,降幅再次增大,分别为27.5%~29.5%和40.7% 和48.6%。复水后气孔导度的恢复与胁迫强度和持续时间有关。复水7 d后,高度水分胁迫7 d 组在光量子通量密度高于1000 μmol·m-2·s-1时叶片gsw比轻度水分胁迫具有更强的恢复能力,gsw较CK高38.3%~61.2%;当胁迫时间延长到14 d,复水7 d后gsw虽有所升高,但均未达到CK水平;进一步延长到21 d,轻度水分胁迫组gsw仍表现出较强的恢复能力,但高度水分胁迫组gsw则进一步下降,与CK相比降幅达85%以上。
Ci和Ls的变化方向是判断光合过程气孔因素和非气孔因素限制的重要依据,Ci降低和Ls升高表明,气孔导度降低为主要原因,Ci升高和Ls降低表明,以非气孔限制为主[27]。图5是孕穗开花期水分胁迫7 d和21 d处理组的Ci和Ls随光量子通量密度的变化。由图5可知,水分胁迫处理7 d后,800 μmol·m-2·s-1光量子通量密度以下,随光强升高Ls升高Ci下降,800 μmol·m-2·s-1光量子通量密度以上,Ls和Ci的变化方向不明显,因此,气孔限制是BH-07和BL-07光合速率降低的主要原因。当胁迫时间延长14 d和21 d时,高光强下随光强升高,Ls降低和Ci升高的倾向率逐渐增加,且随胁迫增强而增大(图略),说明随着胁迫增强和持续时间增加,非气孔因素逐渐成为制约光合过程的主要因素。复水后,水分胁迫7 d和14 d处理组光合过程主要以气孔限制为主(图略),但21 d处理组为明显的非气孔限制。

图4 孕穗开花期叶片气孔导度对光量子通量密度的响应Fig.4 Responses of leaf stomatal conductance to photosynthesis photon flux density at booting and flowering stage

图5 孕穗开花期叶片气孔限制值和胞内CO2浓度对光量子通量密度的响应Fig.5 Responses of stomata limitation and intercellular CO2 concentration to photosynthesis photon flux density at booting and flowering stage
2.3 水分胁迫/复水对灌浆期叶片光合速率和水分利用效率影响
在灌浆期,随水分胁迫增强和持续时间延长,叶片光合速率和水分利用效率变化幅度小于孕穗开花期,且两个发育阶段不同处理天数组间存在较大差异(图6和图7)。灌浆期持续7 d的水分胁迫处理可导致光合速率显著降低,降低幅度低于孕穗开花期相同日数的水分胁迫处理,水分利用效率并未表现出类似孕穗开花期的显著提高。持续时间延长到14 d时,相同光强下,FH-14,FL-14与CK的光合速率无显著差异,FL-14处理组水分利用效率高于CK,FH-14处理组水分利用效率低于CK。总体上,光合速率与水分利用效率变异规律与孕穗开花期基本相同。当水分胁迫持续时间延长到21 d时,虽水分利用效率提高,但光合速率显著降低,且随着胁迫增强降幅增大。
在灌浆期,水分胁迫复水后光合速率和水分利用效率的恢复与胁迫强度和持续时间有关。水分胁迫7 d复水7 d后,水分利用效率提高,与孕穗开花期(图3)表现基本一致;轻度水分胁迫在复水后,光合能力仍具有很强可恢复性,在光量子通量密度大于600 μmol·m-2·s-1时光合速率均显著高于CK,但与孕穗开花期不同的是,FH-07-R并未表现出较强的恢复能力,几乎在所有光强下,其光合速率均显著低于CK(图6)。在灌浆期随着叶龄的增长,谷子叶片光合速率较孕穗开花期有所下降,最大值仅为25.9 μmol·m-2·s-1,且出现较明显的光抑制现象,FH-07-R光合能力恢复有限,可能与高度水分胁迫加速叶片老化,造成器质性的功能损伤有关。随着水分胁迫持续时间延长到14 d及以上,复水7 d后水分利用效率显著提高(图7),但光合速率均未恢复到CK水平(图6)。另外,对比图3和图7可以看到,灌浆期水分胁迫处理组具有比孕穗开花期水分胁迫处理组更高的水分利用效率,这是谷子在灌浆期更耐旱的证据之一。但回勾状的光合/蒸腾速率关系曲线说明(图8),由于叶龄增加,器官老化,非气孔限制缓解困难,在高光强下持续21 d的高度水平胁迫复水后,谷子会以光合速率降低更多为代价,关闭部分气孔避免水分的过分散失,这或可解释为何灌浆期遭遇干旱后产量损失更大(图1)。
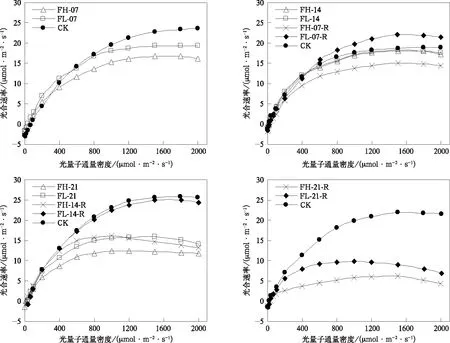
图6 灌浆期叶片光合速率对光量子通量密度的响应Fig.6 Responses of leaf photosynthetic rate to photosynthesis photon flux density at grain filling stage
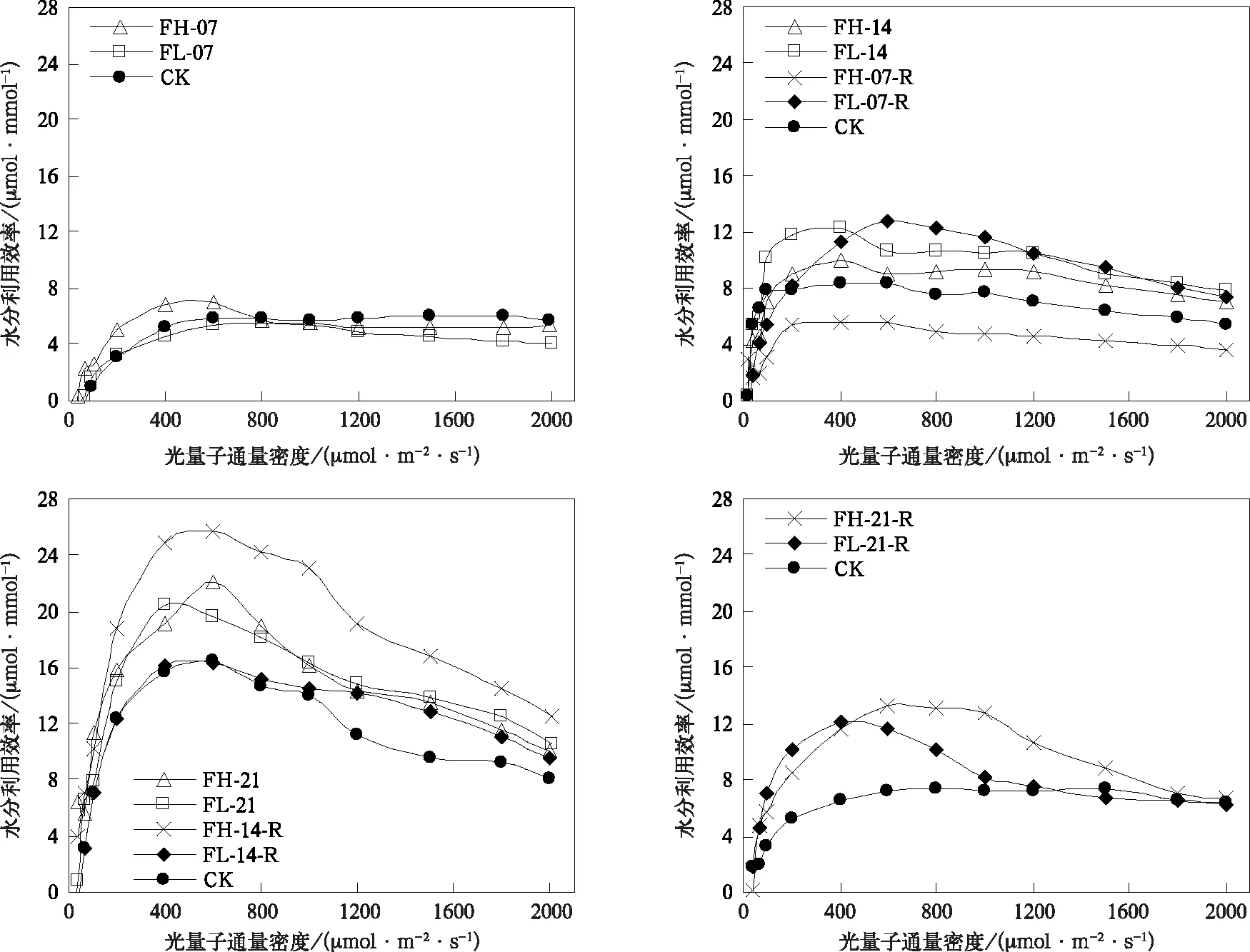
图7 灌浆期水分利用效率对光量子通量密度的响应Fig.7 Responses of water use efficiency to photosynthesis photon flux density at grain filling stage
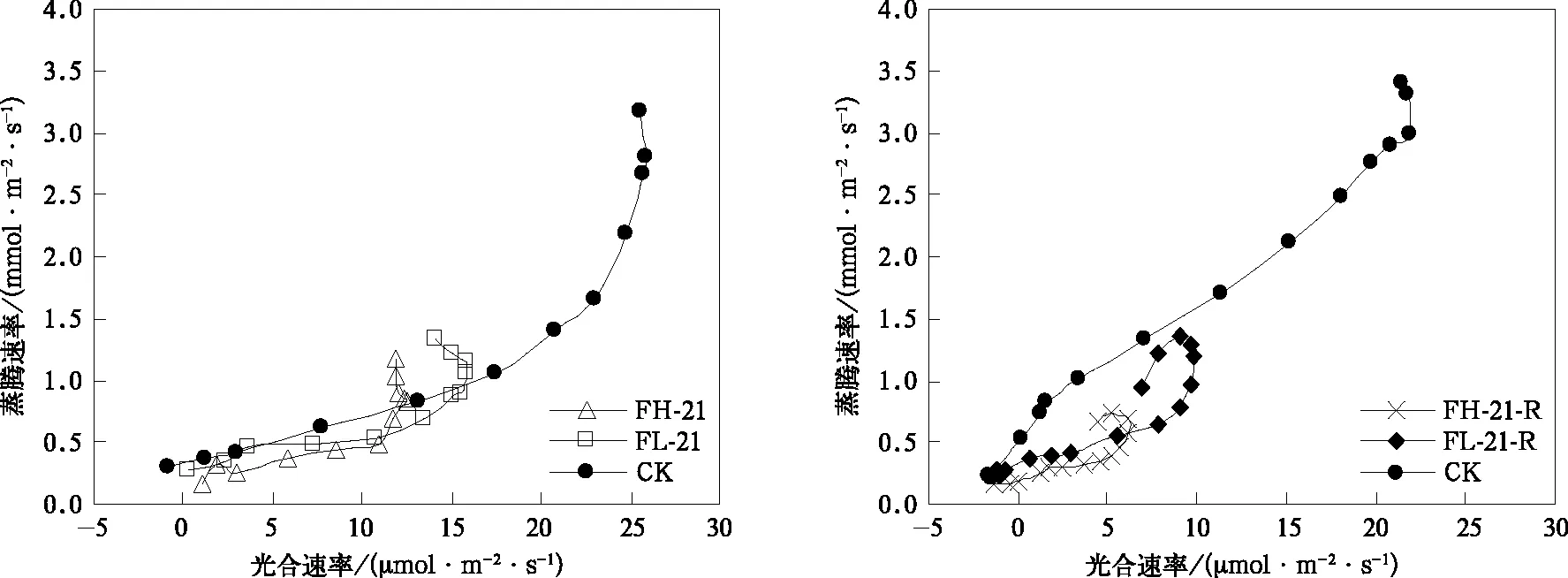
图8 灌浆期叶片光合速率与蒸腾速率的关系Fig.8 Relationships between leaf photosynthetic and transpiration rate at grain filling stage
灌浆期水分胁迫同样可导致叶片气孔导度降低,胁迫越强降幅越大(图略)。与光合速率随持续时间变化类似,灌浆期持续7 d水分胁迫处理组叶片气孔导度下降幅度(图略)小于孕穗开花期(图4),而持续21 d的水分胁迫相反,灌浆期下降幅度(图略)大于孕穗开花期(图4)。复水后,气孔导度的恢复能力与孕穗开花期表现类似。Ls和Ci变化表明:气孔因素是导致FH-07,FL-07,FL-14处理光合速率下降的主要原因,随胁迫时间延长到14 d和21 d,非气孔因素逐渐成为制约光合过程的主要因素,复水后,水分胁迫7 d和14 d处理组的光合能力主要受气孔因素限制,但21 d处理组则表现为明显的非气孔限制,灌浆期不同胁迫强度和持续时间处理组光合速率降低的限制因素与孕穗开花期表现一致。
3 结论与讨论
本研究利用大田控水试验分析不同强度及持续时间的水分胁迫对谷子孕穗开花期和灌浆期光合特性的影响,评价复水后光合性能的恢复状况,阐述气孔及非气孔因素在光合过程中的制约作用,分析不同发育阶段水分胁迫对产量的影响,得到如下结论:
1) 谷子遭遇水分胁迫,会导致光合速率下降和水分利用效率提高,其变化幅度与胁迫强度、持续时间及发育阶段有关。随胁迫增强和持续时间延长,光合速率下降越明显,轻度水分胁迫下,谷子表现出更积极的水分利用策略。复水后,光合作用可产生补偿效应,光合性能有所恢复,孕穗开花期14 d以内的水分胁迫和灌浆期7 d以内的轻度水分胁迫,复水后能较快恢复正常。
2) 在轻度水分胁迫和持续时间较短的高度水分胁迫下,光合过程以气孔限制为主,随着水分胁迫强度的增强和持续时间的延长,非气孔因素逐渐成为光合作用降低的主要原因。在相同强度和持续时间情况下,灌浆期的非气孔限制强于孕穗开花期。
3) 水分胁迫/复水处理后,谷子叶片尺度光合速率变异规律与最终产量变化具有一致性。与孕穗开花期相比,产量对灌浆期水分胁迫更敏感。
作物在遭受水分胁迫后光合性能明显下降[29],且随胁迫程度加重,下降愈大[30-32],但复水后能够产生补偿效应[33-34]。本研究表明:孕穗开花期和灌浆期水分胁迫均显著降低谷子叶片的光合速率,且降低的幅度与水分胁迫时期、强度和持续时间有关,与小麦[35]、黑麦[36]的研究结论一致。复水后谷子叶片光合性能可产生一定补偿效应,补偿效应的大小与水分胁迫的强度和持续时间有关。孕穗开花期14 d以内的水分胁迫和灌浆期7 d以内的轻度水分胁迫,复水后能较快恢复正常,这可能是水分胁迫尚未严重损及光合器官及其生理机能,因此具有较强的恢复能力;而孕穗开花期21 d的水分胁迫、灌浆期14 d及以上的轻度水分胁迫和7 d及以上的高度水分胁迫可能导致谷子光合器官结构和功能受到一定程度的破坏,复水后光合能力的恢复较弱。
干旱情况下,作物水分利用效率提高是其响应环境变化的主要特征[13-14]。本研究表明:谷子在水分胁迫下,可通过提高水分利用效率减少光合产物损失。谷子在遭遇7 d和14 d的水分胁迫时,轻度水分胁迫处理组具有更高的水分利用效率,说明轻度水分胁迫处理组比高度水分胁迫处理组具有更积极的水分利用策略,对干旱的响应迅速以适应干旱;当持续时间延长到21 d时,高度水分胁迫处理组具有更高的水分利用效率则是出于“保命”需求。复水后,水分利用效率仍能保持在较高水平,且灌浆期水分胁迫处理组具有比孕穗开花期更高的水分利用效率,这是谷子在灌浆期更耐旱的证据之一。但回勾状的光合/蒸腾速率关系曲线说明,叶龄增加和器官老化导致非气孔限制缓解困难,在高光强下持续21 d 的高度水平胁迫即使在复水后,谷子仍会关闭部分气孔避免水分的过分散失,以光合速率降低为代价,这可解释灌浆期遭遇干旱后产量损失更大。
气孔对植物光合作用和蒸腾作用的平衡具有调节作用,在植物适应环境变化和环境压力中起关键作用[27,37-39]。谷子在水分缺乏时,关闭部分气孔可以减少叶片蒸腾耗水,从而有利于提高水分利用率,是谷子为抵抗干旱胁迫,对干旱的自我适应[40-41]。本研究表明:在轻度水分胁迫和持续时间不长的高度水分胁迫下,光合过程以气孔限制为主,随着水分胁迫强度的增强和持续时间的延长,非气孔因素逐渐成为光合作用降低的主要原因;灌浆期的非气孔限制略强于孕穗开花期,这可能与叶片老化,灌浆期水分胁迫后叶片光合器官结构和功能受到伤害有关[25]。
光合过程是产量形成的基础和保证。作为我国传统的旱粮作物之一,谷子具有较强的抗旱能力,但干旱仍然会对其光合过程及产量产生不利影响[42]。本研究不同发育阶段水分胁迫/复水处理,其光合速率的变异规律与产量变化具有一致性。根据本试验的产量构成数据发现,孕穗开花期的产量构成主要表现为穗粒数的减少和千粒重的增加,在水分胁迫/复水光合作用变化不大的情况下,二者对产量影响可部分抵消,可能是造成该时期产量对水分胁迫的响应弱于灌浆期的原因;灌浆期产量的降低主要表现为千粒重的下降,光合速率降低直接导致充实不足,使得该时期产量与光合速率一致性更高。
