沙门氏菌生物菌膜形成的分子调控研究进展
2021-07-29王虎虎何淑雯胡海静邵良婷徐幸莲
王虎虎,何淑雯,胡海静,白 云,邵良婷,徐幸莲
(南京农业大学 肉品加工与质量控制教育部重点实验室,江苏 南京 210095)
食源性致病菌是危害公众健康和食品安全的主要因素[1],全球范围内的污染调查结果显示,在所有食源性致病菌中,沙门氏菌每年引发的食物中毒事件数量位居首位[2-3]。污染溯源结果显示大量的微生物污染主要来源于交叉污染,存活于食品加工设备表面的黏附细菌和生物菌膜是引起交叉污染的源头和传播中心[4-5]。生物菌膜是细菌黏附行为最为突出的表现,也被称为生物膜或生物被膜,指菌体吸附于固体材料表面(如食品加工设备中的不锈钢、塑料和玻璃等),并通过增殖、分泌胞外基质而形成的具有一定空间结构的细菌群体[6]。相比于浮游态,黏附菌体对食品消毒剂的抵抗性提高了成百上千倍,并可促进毒力和耐药基因在细菌种属间的水平转移,对食品安全具有重大危害。在生物菌膜形成过程中,初始黏附和生物菌膜成熟是最为重要的两个阶段,初始黏附与菌体的黏附潜能密切相关,而生物菌膜成熟直接由胞外多聚物(extracellular polymeric substances,EPS)决定,这两个过程均受相关基因通路的调控,因此初始黏附和EPS及其调控通路是研究生物菌膜的主要聚焦点。本文在阐述黏附辅助体(菌毛和鞭毛)和EPS对生物菌膜形成的具体作用基础上,重点论述了mRNA和非编码小RNA(non-coding small RNA,sRNAs)对生物菌膜形成的调控途径、机制和作用模式。
1 生物菌膜的mRNA调控通路
在生物菌膜形成的初始阶段,菌体结构(如鞭毛和菌毛)在靠近并附着于接触物表面的过程中起主要作用。鞭毛、I型菌毛和IV型菌毛对菌体与各类接触物表面的初始相互作用至关重要,能使细菌沿着接触物表面移动,从而促进生物菌膜的生长和扩散[7-8](图1)。在生物菌膜成熟期,主要靠EPS包裹菌体并维持三维网状结构,细菌从浮游态转化为黏附态的过程中,控制相关菌体结构和EPS合成的mRNA调控通路发挥了重要作用,直接决定生物菌膜形成能力。

图1 生物菌膜形成模型[8]Fig. 1 Model for biofilm formation[8]
1.1 鞭毛调控
浮游菌首先与接触物表面接触实现初始黏附,进而形成生物菌膜,此过程主要由鞭毛介导的移动性主导,菌体可通过鞭毛的牵引作用克服排斥力后移动至接触物表面[9],这是细菌黏附的先决条件。鞭毛由基体、鞭毛钩和鞭毛丝3 个部分组成[10]。根据鼠伤寒沙门氏菌的鞭毛调节基因模型,在转录层面的鞭毛操纵子分为3 类,第1类仅包含flhDC1 个操纵子,第2类包含7 个操纵子(flgA、flgB、flhB、fliA、fliE、fliF和fliL),均受第1类操纵子的正调控,第3类包含至少5 个操纵子(flgK、tar、motA、fliC和fliD)。鞭毛操纵子的顺序表达与鞭毛结构的组装过程相关联,所有与钩体组装有关的基因都属于第2类,而与丝体组装有关的基因都属于第3类。以flhDC为核心的鞭毛形成通路[11]在细菌指数生长期被激活,flhDC激活由fliA编码的σF因子以及fliA下游基因,这些元件共同调控菌体鞭毛的形成,并决定着菌体的移动性能;同时,σF因子调控着由YhjH介导的YcgR蛋白表达,以此降低YcgR对鞭毛形成的抑制作用。尽管随着黏附进程的推进,与鞭毛形成的关联基因被显著下调,但不可否认的是鞭毛在菌体初始黏附阶段和决定菌体移动性能方面发挥着关键作用。
1.2 菌毛调控
I型菌毛是肠杆菌科中常见的一种黏附细胞器,主要负责与宿主细胞的首次接触以及宿主与病原体间的相互作用[12]。I型菌毛可通过I型黏附素FimH与接触物表面之间的相互作用,非特异性地附着于多种接触物表面,在生物菌膜形成的早期阶段发挥重要作用[13-14]。肠炎沙门氏菌的I型菌毛参与菌体对不锈钢或聚四氟乙烯等表面的黏附[15]。IV型菌毛是常见的细菌表面附属结构,IV型菌毛通过聚合而伸长,通过解聚而回缩,其长度是动态变化的,该特性对于细菌在接触物表面的定植具有重要作用。在回缩过程中,当IV型菌毛附着到接触面时,会向该接触面施加非常大的力,该力可以触发细菌的蹭行移动,实现细菌在接触物表面的移动[16]。同时,IV型菌毛能够通过稳定菌体与接触物表面的相互作用和对抗菌体-菌体间的静电斥力促进微菌落和细菌单层形成[9],为生物菌膜的形成奠定基础。肠道沙门氏菌中,仅在严格适应人类的伤寒血清型中检测到IV型菌毛,组装其所必需的11 个基因在沙门氏菌“致病岛”上的菌毛操纵子中均存在,这些基因是该菌黏附至人肠细胞所必需的[17]。
1.3 EPS调控
胞外基质是生物菌膜的另一重要组分,主要由EPS构成,根据具体功能,不同的EPS可被划分为结构性、吸附性、表面活性和信息性等多种类型。EPS在生物菌膜形成中的主要作用包括但不限于[18-19]:1)浮游菌与基质接触时,通过分泌EPS可实现迅速而牢固的黏附;2)局部的EPS分泌会引起细菌的滑动,允许其趋向更佳的生长条件或与其他微生物获得联系;3)细菌在胞外分泌大量的EPS,有利于生物菌膜稳定;4)生物菌膜成熟后,EPS的持续产生能将部分细胞推离接触物表面,使其进入流动相并脱离生物菌膜。
沙门氏菌EPS的主要成分有纤维素、淀粉样卷曲纤维、荚膜异多糖酸等。其中,纤维素(αβ-1-4-D-葡萄糖聚合物)是一种结构性EPS,能够与其他胞外组分交织形成三维结构,促进细胞间相互作用,并保护细胞免受不利环境的侵害。它的合成需要bcsABZC-bcsEFG操纵子编码的基因参与。卷曲纤维蛋白是一种淀粉样蛋白物质,其在大肠杆菌中的生物合成已被广泛地报道[20],同源操纵子在沙门氏菌属中已被鉴定出,并被命名为agfBA和agfDEFG;肠炎沙门氏菌能够产生多种类型的卷曲纤维蛋白,包括SEF14、SEF17、SEF18和SEF21,它们具有黏附到特氟龙和不锈钢等多种接触物表面的能力[21]。荚膜异多糖酸是一种复杂的支链聚合物,其合成需要wca基因座中携带的19 个基因;无法合成荚膜异多糖酸的突变体可以形成一到两层细胞厚的致密层,但无法构建更复杂的多层生物菌膜[22]。在黏附基质形成通路中,csgD是生物菌膜形成的核心调控元件,其在细菌指数生长末期和稳定期大量转录表达,并受温度、氧分压、高渗透压和酸度等外源应激因子的调控[23],这也是菌体能够快速响应环境变化并适应存活的分子基础;csgD几乎决定了所有黏附体EPS的形成和分泌,它通过csgB、adrA、bapA和yihW等基因的调控实现对菌毛、菌体纤维素和表面黏性蛋白等黏附因子的合成控制,综合调控黏附聚集体的最终形成和稳定性。
1.4 其他调控通路
RpoS(σS)是一种细菌系统调控元件,调控着500多种与稳定期生长和胁迫响应关联的基因,可为生物菌膜中的菌体提供强大的保护。沙门氏菌生物菌膜形成过程中,超过25%的与rpoS调控有关的基因表达都会显著上调[24-25]。同时,生长稳定期菌体内csgD的表达及其对csgB和adrA的调控也会显著依赖于rpoS,同时rpoS可以激活与移动性能有关的通路进而对生物菌膜产生影响。
c-di-GMP是细菌体内一类重要的第二信使,它是三磷酸鸟苷在二鸟苷酸环化酶、磷酸二酯酶等物质生化反应下的最终产物,该信号分子通过与关联蛋白(CsgD、BcsA和AdrA等)结合改变活性方式,促进菌毛和菌体纤维素的形成或抑制菌体移动性从而参与生物菌膜的形成调控;已证实菌体纤维素的合成完全依赖于体内c-di-GMP水平,且菌体对该信号分子的自我利用没有形成来源偏好性[26]。
除上述通路外,沙门氏菌内还存在phoP/Q、csrA-barA/sirA、csr和Rcs等与菌体黏附行为有关的其他辅助调控通路[26-28],这些通路通过影响菌体的黏附潜能、移动性能、外膜蛋白、抗应激能力、二级代谢、EPS和细菌趋向性等途径调控生物菌膜形成。
2 sRNA对生物菌膜的调控效应
2.1 sRNA简介
sRNAs是近年来在原核生物中发现的一类新型转录后调控因子,广泛存在于沙门氏菌等食源性致病菌中。sRNA的长度介于50~500 nt之间,在基因组中被转录但不翻译蛋白质,其转录通常始于可折叠形成稳定茎环结构的基因序列,终止于Rho不依赖的转录终止子[29]。sRNAs主要通过两种方式发挥调控作用[30]:1)通过“碱基互补配对”的方式抑制靶mRNA的翻译或降解,该过程通常需要伴侣蛋白Hfq的参与协助[31],这也是sRNA最主要的调控方式;2)通过模仿核酸结构调控靶标蛋白质活性,sRNAs竞争性结合调控蛋白(RNA结合蛋白)的靶标mRNA,抑制靶标蛋白的功能发挥。sRNAs对细菌生理代谢功能的调控已成为当前的研究热点之一,部分研究机构建立专门的sRNAs资源库平台,供全球学者研究共享,如Rfam、sRNATarBase和BSRD(bacterial small regulatory RNA database)等。
沙门氏菌是学者们研究细菌sRNAs的模式菌株之一,尤其是鼠伤寒血清型,目前针对该血清型的研究最为集中和系统。研究人员利用微矩阵、生物信息学预测、高通量测序等技术手段,现已在沙门氏菌中发现了140多个sRNAs,有些sRNAs在沙门氏菌属内保守,如IsrL和InvR;有些是某种血清型特有,如sRNA Stnc1400只存在于鼠伤寒和乙型副伤寒血清型的某些菌株中[32]。沙门氏菌的sRNAs参与了菌体生长存活、毒性侵袭、黏附和耐受应激等多种代谢调控[33-35]。
2.2 sRNA对移动性的调控
细菌的移动性在生物菌膜形成的早期发挥重要作用[10],Kearns将细菌移动划分为丛动、泳动和蹭行等多种方式[36]。上述移动方式主要由鞭毛和菌毛等器官提供动力。目前,已有诸多研究表明sRNA参与了沙门氏菌移动性的调控。在鼠伤寒沙门氏菌中,sRNA SroC能够通过下调flhBAE和fliE基因的表达以调节鞭毛的合成,且SroC过表达的野生株呈现出了非鞭毛表现型[37]。Ryan等发现sRNA DsrA的缺失降低了鼠伤寒沙门氏菌的移动性能,并指出DsrA可能具有上调鞭毛结构蛋白基因fliC和fljB的作用[35]。同时研究发现参与运动性和趋化性的3 个基因(flgJ、cheY和fliF)的转录受sRNA RyhB同源物RyhB-2下调[38]。另外sRNA McaS可通过与flhD5’-UTR中的两个区域结合而直接激活鞭毛合成的主要转录调节因子flhDCmRNA,从而导致细菌运动性的增强[39]。国内学者陆仁飞等发现鼠伤寒沙门氏菌sRNA S2959通过上调flhD、fliA和fljB的mRNA水平促进鞭毛的表达,进一步提高细菌的移动和菌膜形成能力[40]。
在其他细菌中也发现sRNA参与菌体的移动性能调控,在大肠杆菌中发掘出9 种能够同时抑制泳动和丛动的sRNA[41],研究发现GlmY可导致泳动能力的大幅下降,但对丛动的影响较为轻微;与之相反,SgrS(一种抑制葡萄糖转运的sRNA)能显著抑制丛动,但对泳动作用甚微。此外,研究表明蹭行运动部分取决于prrF基因座,揭示了sRNA PrrF与运动性之间的新型联系[42]。
2.3 sRNA对EPS的调控
csgD是控制生物菌膜形成的核心转录因子,能够诱导产生卷曲菌毛和纤维素以及调控c-di-GMP所需的csgBAC操纵子的表达。csgD的表达在转录后层面至少受到5 个Hfq依赖型sRNA的调控,分别为OmrA、OmrB、GcvB、McaS和RprA[43],均能与csgDmRNA的5’非编码区(长约150 nt)发生碱基互补配对,通过封闭核糖体结合位点、干扰翻译起始因子而发挥阻遏作用。这些sRNA分别属于不同的调节子,并且在不同的生长条件下表达。
在响应高渗透压下,OmrA/OmrB的表达由EnvZ-OmpR进行调控,在适合细菌浮游生长的条件下,OmrA/OmrB的过表达可能导致csgD的下调,进而造成卷曲菌毛的减少。sRNA GcvB是氨基酸转运的转录后总调节因子,由参与调节甘氨酸裂解系统的两个主要转录因子GcvA和GcvR进行调节,能够在响应氨基酸可获得性时抑制csgD的表达。McaS是肠杆菌中一种重要的sRNA,由碳源的质量和生长速率激活[43-44],可通过结合csgD和flhCDmRNA并反向对其调控而不利于生物菌膜形成:在McaS缺失的情况下,csgD会发生上调,而McaS表达量的增加会激活鞭毛主调节子flhDC的合成。依赖McaS的生物菌膜调控较为复杂,因为该sRNA还能够激活聚-N-乙酰氨基葡萄糖(一种广泛存在于生物菌膜胞外基质中的多糖),导致形成独立于CsgD途径的生物菌膜[45]。sRNA RprA在应激时被激活,能够正向调控一般应激反应σS,一旦细胞转变为黏附态,RpoS就会激活csgD的表达和卷曲纤维蛋白的生物合成,而RprA则会通过二鸟苷酸环化酶YdaM抑制csgD表达。此外,RprA能够在卷曲纤维蛋白存在情况下调节荚膜异多糖酸和纤维素的合成,为了平衡响应特定环境刺激所必需的EPS组分的表达[45]。
2.4 sRNA对其他核心通路的调控
在σS控制的一般应激反应的诱导中,DsrA、ArcZ和RprA3 种sRNA起到了重要作用[25,46-49]。其中,DsrA决定了当细胞处于低温(<25 ℃)或暴露于高渗透压时RpoS的累积及其活性的保持。RprA由Rcs激活,能直接抑制包括鞭毛合成基因在内的其他基因;渗透压升高也可诱导Rcs系统,在DsrA缺失的情况下,RprA能够在渗透压冲击条件下激活rpoS的翻译;并且,RprA激活的rpoS翻译有助于确保生物菌膜成熟过程中RpoS的正确定时表达。ArcZ由双组分系统ArcB/ArcA负调控,ArcZ从最初120 nt转录本的3’末端被加工成56 nt的RNA,56 nt的ArcZ可与rpoS发夹配对,采用与DsrA和RprA类似的方式激活rpoS翻译;在有氧条件下,ArcZ表达良好,但在无氧条件下,ArcA抑制ArcZ表达,从而间接地下调rpoS翻译。
研究表明,c-di-GMP的水平直接或间接地受到sRNA的调节。在肠杆菌中对c-di-GMP水平的间接调节已确定与生物膜调节因子CsgD相关。CsgD能够激活鸟苷酸环化酶AdrA,并促进c-di-GMP的合成。另一方面,CsgD反过来通过鸟苷酸环化酶YdaM受到c-di-GMP的作用,从而对卷曲菌毛的表达进行调控。CsgD和YdaM均受sRNA RprA负调控[49],RprA产生的负前馈环能直接下调CsgD和YdaM所控制的基因,确保c-di-GMP维持在低水平。群体感应反应所需的合酶LuxS参与了鼠伤寒沙门氏菌生物菌膜的形成过程,luxS突变株的生物菌膜表型取决于sRNA MicA,且该sRNA的平衡浓度对于生物菌膜形成至关重要,但其具体作用机制有待进一步研究[50]。
CsrB和CsrC是两个经过充分研究的sRNA,能够通过控制游离CsrA/RsmA蛋白的水平以触发生物菌膜形成。近年来,对CsrB和CsrC作为细菌基因表达调控因子的研究大幅增加;CsrB/CsrC和CsrA的直系同源物已在许多其他伽玛变形杆菌中鉴定出来,包括肠炎沙门氏菌、假单胞菌(RsmX/RsmY/RsmZ和RsmA)、弧菌(CsB、CsrC/CsrD和CsrA)和耶尔森菌(CsrB/CsrC和CsrA)等[45]。
2.5 sRNA对生物菌膜的调控模式
黏附行为是沙门氏菌应对外源环境胁迫而常采取的一种自我保护性行为,同时也是发挥菌体侵袭毒性的先决基础。以沙门氏菌和大肠杆菌为代表的肠杆菌科sRNAs主要通过以下4 种模式发挥其对黏附行为的调控作用(图2)。
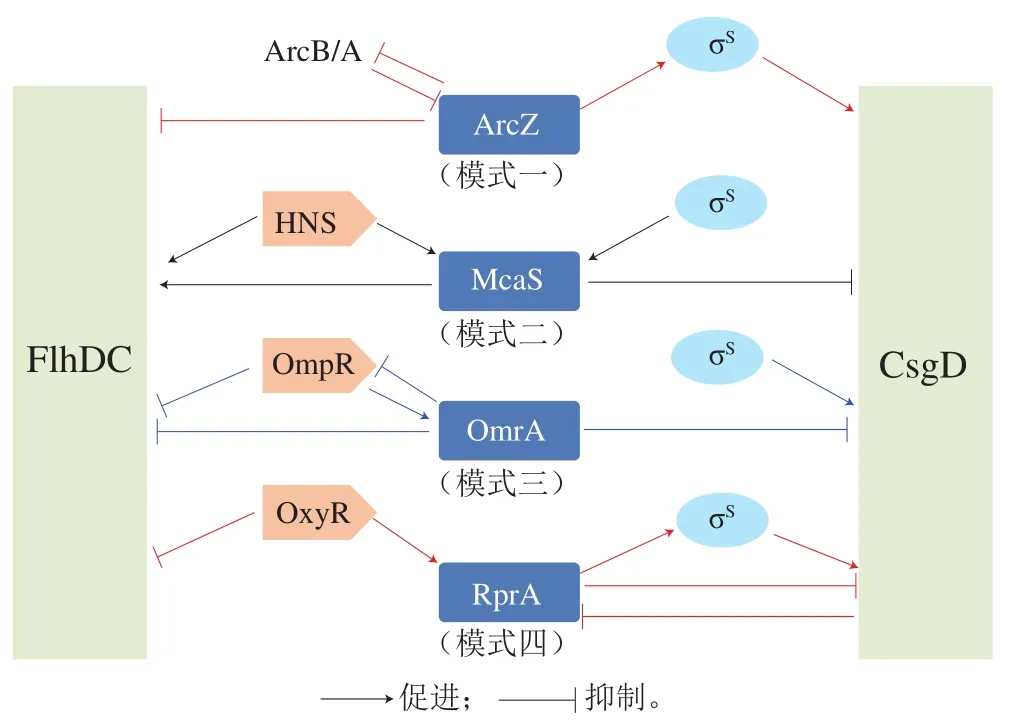
图2 sRNAs对黏附行为的调控模式[11]Fig. 2 Regulation patterns of sRNAs on Salmonella adhesion[11]
模式一:抑制鞭毛产生、促进生物菌膜形成。该模式是sRNAs调控黏附行为的经典模式,ArcZ是该模式下研究的最为清楚也最有代表性的sRNAs[51],沙门氏菌基因组中16%的基因表达都与ArcZ有关,其可通过碱基配对的方式直接与flhDC5’-UTR发生结合从而抑制鞭毛的形成;在ArcB/ArcA双组分系统的参与下,ArcZ可激活对csgD表达具有促进作用的rpoS转录[52-53],并显著降低菌体泳动能力。此外,DsrA和GadY也具有此类模式的调控功能。模式二:促进鞭毛产生、抑制生物菌膜形成。该模式是沙门氏菌sRNAs调控黏附行为的一种潜在模式,McaS是细菌碳循环调控网络中的重要组成,其可通过结合flhD5’-UTR的序列直接激活flhDC,并通过与csgD的5’端部分结合,降低csgD表达[11],同时诱导csgD发生自我降解[44],在最适生长的37 ℃下该sRNA的调控作用最为显著[25]。模式三:双重抑制模式。该模式下sRNAs对鞭毛表达和生物菌膜形成具有双重抑制作用,代表性sRNAs有OmrA/B,其可直接抑制由OmpR介导的鞭毛合成和FlgM的形成[54],并降低卷曲纤维蛋白菌毛和菌体纤维素的形成量[55-56];菌体生长环境中的应激条件可诱导OmrA/B高效转录,在高渗透压应激下此类调控模式尤为显著。模式四:多重效应模式。此状态下的代表性sRNAs有RprA,其过量转录可直接下调csgD。特定条件下,RprA可作为激活因子直接诱导σS介导的调控因子,并通过抑制YdaM表达进而间接干预csgD的转录;当csgD足量表达时,RprA也是其潜在的结合目标,进而降低RprA与其他调控mRNA的结合效率[49]。
3 结 语
沙门氏菌生物菌膜广泛存在于食品工业中,揭示其形成过程和影响因素是研发针对性控制措施的先决基础。本文在总结前人研究发现的基础上,从决定细菌生物菌膜形成过程中最为重要的两个因素——黏附辅助体(菌毛和鞭毛)和EPS入手,系统论述了其在黏附过程中的详细作用,及其在mRNA转录层面上以csgD、flhDC、rpoS等为核心的调控通路,并重点在转录后层面上阐述了sRNA对鞭毛组装和胞外分泌物调控的作用模式。纵观目前的研究进展以及生物菌膜与食品工业的关系,在该领域今后的研究中需重点关注:1)阐明特定食品加工和贮藏环境对沙门氏菌生物菌膜形成的影响,尤其是在外源物理和化学应激条件下的形成过程;2)借助现代组学技术,在转录、转录后等水平上挖掘更多、更详尽的生物菌膜调控通路,尤其是针对全球范围内沙门氏菌污染高、毒性强的血清型;3)解析生物菌膜中关键组分的相互作用及其对黏附体空间特征结构的维持效应;4)利用现代微观成像和示踪技术可视化解析沙门氏菌的黏附与生物菌膜形成过程。
