新疆原类梅属地衣的初步研究
2021-07-28阿布地力木阿布来提范胜男艾木拉古丽达迪汗热衣木马木提
阿布地力木·阿布来提,范胜男,艾木拉古丽·达迪汗,热衣木·马木提
(新疆大学 生命科学与技术学院,乌鲁木齐 830046)
地衣是由藻类和真菌(或蓝细菌)构成的共生复合体,真菌通过菌丝吸收水分和矿物质供给藻类,藻类通过光合作用为菌丝提供营养成分,是一种典型的共生生物[1]。原类梅属(ProtoparmeliopsisM. Choisy)隶属于真菌界(Fungi)子囊菌门(Ascomycota),子囊菌纲(Ascomycetes)茶渍目(Lecanorales)茶渍科(Lecanoraceae)[2]。世界范围内原类梅属有48种[3],其中中国已报道5种[4-5]。
原类梅属由M. Choisy建立于1929年,并将石墙原类梅(ProtoparmeliopsismuralisM. Choisy)指定为该属的模式种[6],由于没有被广泛接受的分类和命名依据,Poelt[7]将原类梅属被列入到裂片状茶渍亚属(L.Subg.Placodium)。Poelt等[8-9],Ryan BD等[10-11]对具有裂片状地衣体的茶渍科种类进行了区域性订正研究并编制了检索表。Arup U等[12]以ITS和5.8S分子数据为基础对裂片状茶渍亚属地衣进行系统分类学研究,结果表明,裂片状茶渍亚属地衣的地衣体形态并不能准确地反映其系统发育关系。2000年Arup等[13]证实了原类梅属在属水平的地位,并提供了该属的第一批分子数据。Santesson在《瑞典和挪威的地衣和地衣真菌》[14]一书中发表了原类梅属的4个种类。Kondratyuk等[15-17]报道了原类梅属地衣27个种类。Kondratyuk等[18]完善了原类梅属地衣的分子数据,并提出了该属同脐鳞属(Rhizoplaca)和茶渍属(Lecanora)一样为多源性类群;Miadlikowska等[19],赵欣等[20]、Kondratyuk等[21]和Katarzyna等[22]的系统学研究表明,原类梅属地衣属于茶渍科,并有较高支持率。
原类梅属地衣是壳状地衣,地衣体通常呈脐状或裂片状,黄绿色至枯草黄色,子囊盘茶渍型,土色至淡黄褐色,有时变黑,茶渍型子囊,子囊孢子为无色,透明。该属的物种分布地区主要在半干旱地区,特别是在海拔较高的地区,基物多为硅质岩或土壤[20]。
本研究对采自新疆的原类梅属地衣标本通过形态解剖学、地衣化学及分子生物学等多种方法进行了分类,发现了1个中国新记录种,并对新记录种地衣进行了详细描述,对其系统位置进行了分子系统发育分析。另外,提供了该地区常见的原类梅属地衣其他种类外部形态和内部构造特征、生境、彩色图片、采集地和分种检索表。研究结果为新疆地衣区系及中国地衣物种多样性研究提供了科学性资料。
1 材料和方法
1.1 材 料
研究材料为2011-2016年采自新疆的原类梅属地衣馆藏标本,现保存于新疆大学中国西北干旱地衣研究中心地衣标本室(XJU-NALH)。
1.2 方 法
1.2.1 形态解剖学观察及化学成分检测通过形态学研究方法记录原类梅属地衣体颜色,下地衣体颜色及分布区域,裂片形态,子囊盘类型,子囊盘大小,子囊盘盘缘类型等宏观特征。通过解剖学研究方法记录原类梅属地衣子囊盘上子实层颜色厚度,子实层颜色和厚度,下子实层颜色及厚度,囊盘被厚度及颜色,子囊孢子大小,颜色及形态等微观特征。使用SZM45体视解剖镜观察地衣形态特征,使用Nikon DS-Fi2显微镜和Canon PC1200数码相机进行显微观察及拍照。
通过地衣化学方法记录原类梅属地衣体与K溶液,C溶液,KC溶液以及I溶液的显色反应。通过显色反应(CT)和薄层层析(TLC)等方法鉴定地衣的次生代谢产物[23]。
1.2.2 分子系统发育分析基因组DNA提取使用了真菌基因组DNA提取试剂盒。PCR扩增rDNA-ITS位点,采用真菌通用引物ITS 1F和ITS 4[24-25]。扩增是在25 μL反应体系中进行,其中包括9.5 μL ddH2O,12.5 μL 2× PCR Master Mix,1 μL DNA模板以及各个引物1 μL (10 μmol/L)。扩增ITS 的反应条件为:94 ℃预变性5 min,94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸60 s,5个循环;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸60 s,30个循环;72 ℃延伸5 min。PCR扩增产物的测序工作由上海生工公司完成。
序列拼接和整理使用DNASTAR-seqman软件,使用NCBI 中的Blast工具查找序列相似性最高的序列下载(本研究中下载了16条序列)。利用MEGA7.0[26]中的Clustal W方法进行多序列比对,然后对序列进行手动校正,保留完整的ITS 1、5.8S和ITS 2的序列。再利用MEGA 7.0软件采用(Neighbor Joining, NJ)以 Tamura-Nei模型(T93)构建系统发育树,用Bootstrap方法检验系统发育树,置信度设为1 000,其余参数设置为默认。该树通过使用Myrioleciscontractula和Myriolecisdispersa作为外类群来生根。利用DNAsp5.0软件分析了序列的单倍型和核酸多样性。参与分析的序列包括自测序列和GenBank中下载的序列自测序列信息见表1。
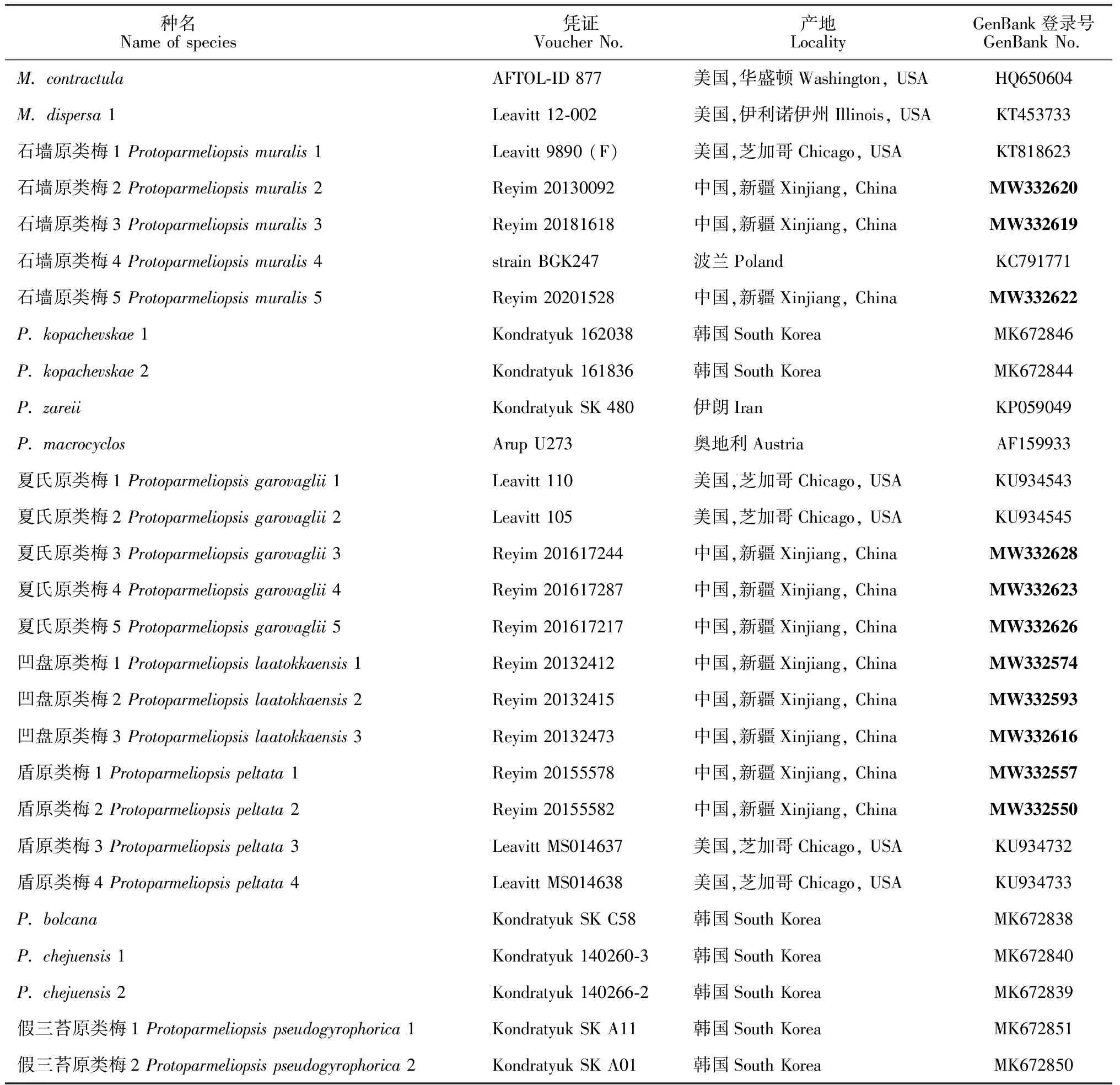
表1 用于系统发育分析的物种及序列信息
2 结果与分析
2.1 原类梅属地衣系统发育分析
通过MEGA软件对序列碱基组成信息,变异位点以及简约信息位点进行分析。ITS序列片段长度507 bp,这些碱基序列中有183个为变异位点,167个为简约位点,284个为保守位点,G十C碱基组成占总含量的57.3%,A十T碱基组成占总含量的42.6%,G+C含量高于A+T含量。G十C含量关系着基因结构的稳定性,其含量越高结构越稳定,因此该研究中ITS基因片段结构较稳定。通过DNAsp5.0软件分析ITS序列的单倍型和核酸多样性,可知28条序列具有18个单倍型,单倍型多样性(haplotype diversity,Hd)为0.958,核酸多样性(nucleotide diversity,Pi)为0.11,二者均较高。
以M.contractula和M.dispersa为外类群,通过邻接法(NJ)对原类梅属地衣ITS基因序列构建系统发育树,通过NJ法分析得出结果所显示的自举值均大于50%。图1显示了NJ法产生的系统发育树拓扑结构,树枝前分别标注了所得的支持率,本研究获得的序列以加黑形式呈现。
由图1系统发育树结果可知,原类梅属地衣被归入为3大分支,其中分支1和分支2形成姊妹群关系,分支1和分支3是非单系群,该系统发育树支持了最近提出的系统发育树结果[22]。在分支1中,石墙原类梅与其他三物种形成了一个具有较高支持率的姊妹群,石墙原类梅自测序列与GenBank数据库中下载的ITS序列聚在一个分支内且支持率为1,证实了该物种的系统学位置。在分支2中,戛氏原类梅自测序列与其他来自GenBank数据库中的序列形成了支持率较高的分支,分子鉴定与形态解剖特征结果一致。
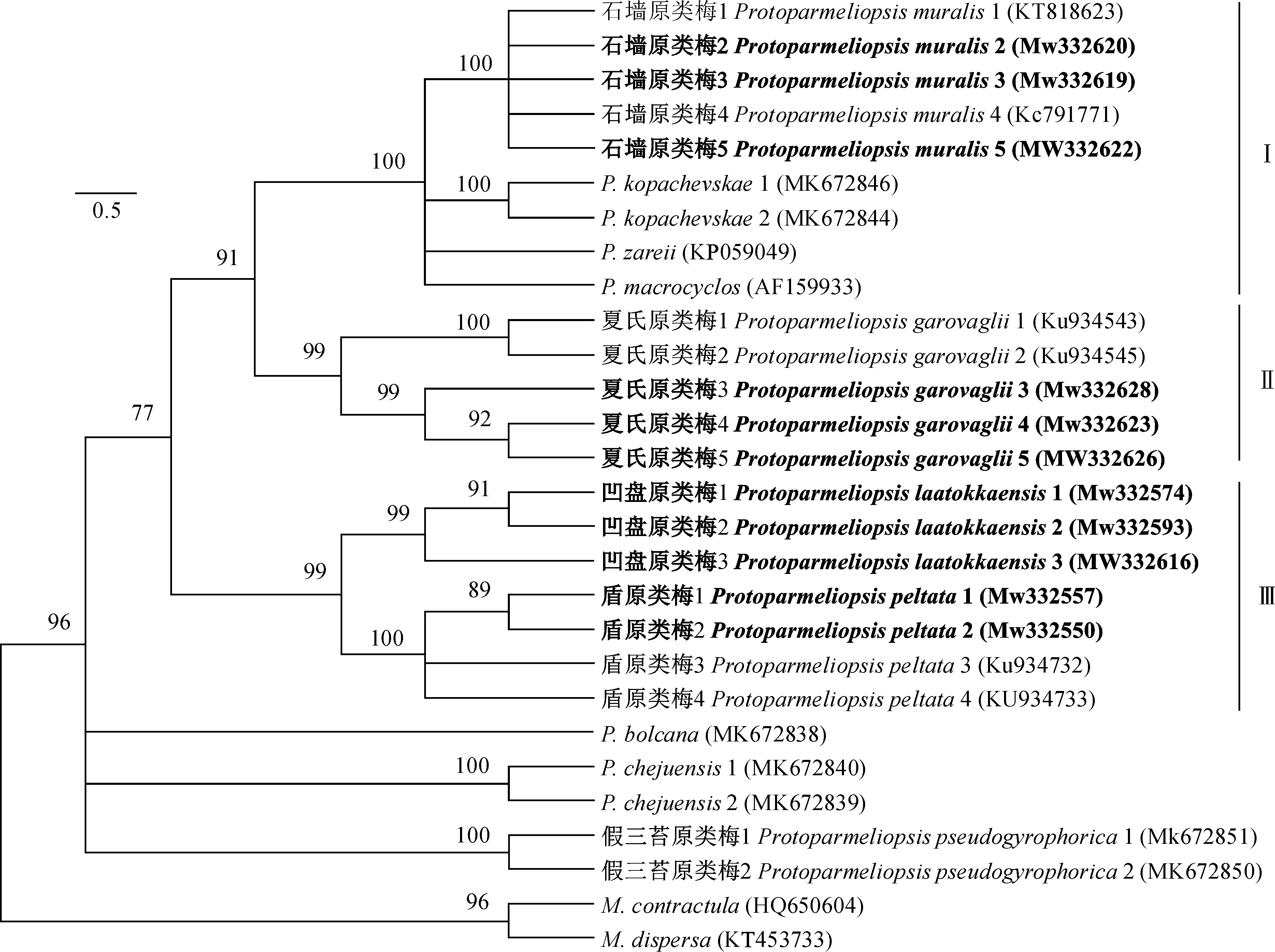
利用邻接法(NJ)构建的系统进化树,数字表示自展指支持值图1 原类梅属ITS序列数据系统发育树Phylogenetic trees constructed using(Neighbor-Joining,NJ) methods,.figures represent the Bootstrap support valueFig.1 Phylogenetic tree of Protoparmeliopsis based on ITS data
2013年Kondratyuk等[17]提出把凹盘原类梅归入到原类梅属,但没有提供其分子数据及系统发育学位置。本研究的分子系统学分析中,凹盘原类梅与盾原类梅聚在同一个分支内(分支3),说明这两个物种亲缘关系比较接近,证实了该物种的系统学位置。在分支3中,盾原类梅自测序列与其他来自GenBank数据库中的序列形成了支持率较高的分支,系统发育树结果与表型特征符合。
2.2 分种描述
2.2.1 凹盘原类梅Protoparmeliopsislaatokkaensis(Räsänen) Moberg & R. Sant. [aslaatokka⊇nsis], in Santesson, Moberg, Nordin, Tønsberg & Vitikainen, Lichen-Forming and Lichenicolous Fungi of Fennoscandia: 269 (2004)(图版Ⅰ, A-F)
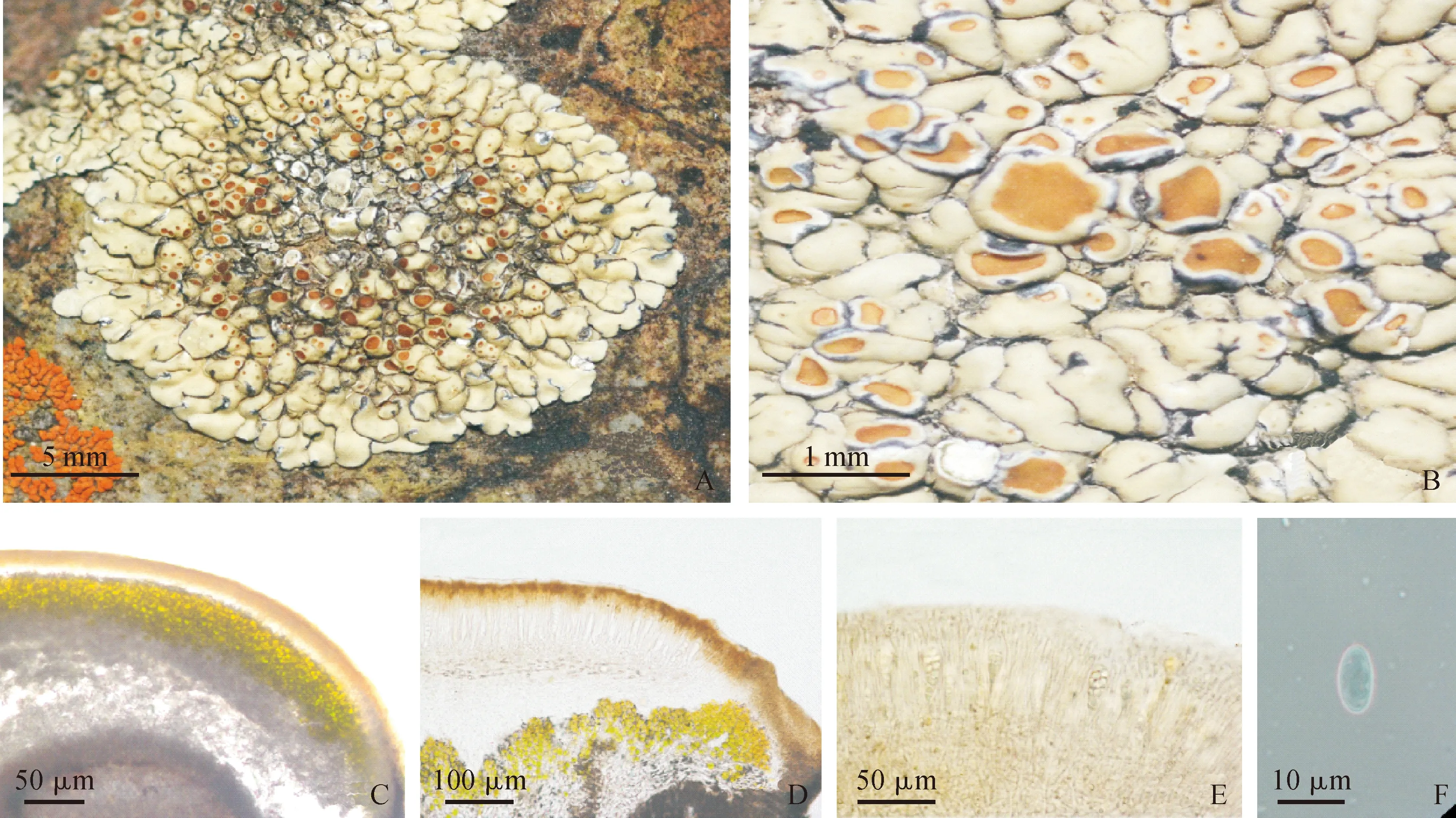
图版Ⅰ 凹盘原类梅(标本号20132490)A-B. 地衣体和子囊盘的外部形态结构;C.子囊盘纵切解剖结构;D.子囊盘纵切解剖KOH溶液处理;E.子囊;F.子囊孢子Plate Ⅰ Photos of Protoparmeliopsis laatokkaensisA-B. Morphological structure of thallus and apothecia; C. Cross section of an apothecium; D. Cross section of an apothecium with KOH treatment; E. Asci; F. Ascospores

新疆原类梅属地衣分种检索表
MycoBank MB366117
地衣体裂片状,黄绿色,网纹状至龟裂状,紧贴于基物,有时凸出,边缘上升,特别不规则,重叠,向边缘成规则形、辐射状;表面光滑,有光泽,边缘有明显相当厚的黑色前地衣体;地衣体顶端小裂片放射状分布,裂片顶端呈半缘至圆齿状,大小介于3~4 mm之间;地衣体上皮层45~75 μm厚;藻层70~100 μm厚,连续或被菌丝较大分离;下皮层黑色或很少发达。
子囊盘茶渍型,子囊盘数量多或在有些标本中很少,浅棕色至棕色,分散至局部聚集生长,凹陷或有时扁平,轮廓呈近圆形至不规则;盘面平整或下沉,无粉霜;盘缘与地衣体同色或稍苍白,与盘面齐平或突起;子囊盘直径约1~2 mm;囊盘被:透明,平行至随机排列的菌丝,约30 μm厚;上子实层:橙棕色,内散有细小颗粒(可溶于K);子实层: 透明的,60~80 μm;侧丝:略膨大,顶端2.5~3 μm宽;下子实层:通常出现有点灰色或棕色,带有油滴;囊层基:中部厚150~300 μm;子囊:棒状,8孢;子囊孢子:通常不发达,透明,单细胞,成熟时椭圆形的11~13 × 4.5~5.5 μm;分生孢子器未见。
显色反应:地衣体K-,C-,KC-,P-;皮层KC+黄色;髓层KC-。
次生代谢产物:zeorin,usnic acid,fatty acids。
基物:岩面生。
分布:欧洲,西亚,北非,北美西部,亚利桑那州东部[27]。中国新纪录种。
引证标本:阿图什乌恰县:玉其塔什草原,20160805,3200,40°13′57″N,74°41′36″E,热衣木·马木提,201617306,201617254,201617287-b;乌鲁木齐县:后峡蓝沟,20091006,2060,N34°00′00″,68°34′37″E,热衣木·马木提,20097231,20098012,20097250;伊宁县:阿吾赞沟,20070703,1450,44°08′28″N,81°40′35″E,热衣木·马木提,20077224,20077241,201617263,20120745;乌鲁木齐县:八一林场,20080411,1864,43°22′43″N,86°47′68″E,热衣木·马木提,20201548,20201547;乌鲁木齐县:一号冰川,20081012,3570,43°06′22″N,86°50′58″E,热衣木·马木提,20201563,20201545,20201546;米泉哈熊沟,20130703,2036,43°48′43″N,87°59′36″E,热衣木·马木提,20201554,20201555,20201556,20201557,20132490,20201558,20201551,20201552,20201562,20201559,20138202,20201564,20132491,20201549;和静县:乔尔玛,20130731,3445,43°28′21″N,84°26′67″E,热衣木·马木提,20132493,20132485,20132508-a,20132508-b;和静县:巴音布鲁克,20130729,2639,43°04′25″N,84°06′48″E,热衣木·马木提,20132418-b,20132380,20201561,20132388-a,20132420-a,20132356,20132425-b,20132388-b,20132448,20132469,20132423,20132449,20132451-a,20132377,20132473,20132475,20132412,20132468,20132419;塔城:托里,20080807,1459,45°49′05″N,83°34′87″E,热衣木·马木提,20081375;和静县:巩乃斯林场,20180816,2150,43°14′47″N,84°40′55″E,热衣木·马木提,20181596-B;国道217-597处,20130731,1418,44°02′89″N,84°41′95″E,热衣木·马木提,20201550。
该种在研究区域内较广分布,与Aspicilia、Xanthoria、Lobothallia、Rhizocarpon和Acarospora种类一起生长。有时寄生在Dimelaenaoreina和一些Aspicilia的种类上。该种最主要的辨别特征是具沉陷入地衣体生长的子囊盘和很明显的黑色下地衣体。该物种与石墙原类梅形态相似,主要区别在于前者形态上裂片较短,子囊盘凹陷,解剖结构没有良好发育的皮层。该种与戛氏原类梅区别在于前者地衣体薄而光滑,裂片短而未凸出,无明显折叠。
2.2.2 戛氏原类梅Protoparmeliopsisgarovaglii(Körb.) Arup, ZhaoXin, Lumbsch, Fungal Diversity, 78(1):301(2015)(图版Ⅱ,G1-G4)
地衣体裂片状,玫瑰花型,具明显的地衣体中心,贴生;地衣体连续但中部多裂至疣状、或瘤状龟裂、或泡状龟裂,边缘裂片状;下地衣体存在,在地衣体裂片之间和边缘末端可以见到,常呈黑橄榄色至淡绿黑色;地衣体边缘连续,表面平坦至强烈不规则的凸出,折成扇状、波状、弯曲,有时形成稍微有规则的沟状裂片,其像植物根茎部的髓射线一样与地衣体中心连接;边缘末端变宽,向内弯曲,呈黑橄榄色至淡绿黑色。地衣体上表面暗淡色,点滴或全部被粉霜,呈淡绿黄色至绿黄色、淡灰黄绿色、绿色、淡绿色,向边缘颜色变淡黄色,常具斑点。上皮层具黄色颗粒(遇K溶解);髓层白色,疏松,蛛网状的,中部空洞,向下颜色变黑;藻胞层不连续,被菌丝分离;下表面淡灰色,网状,呈灰黑色至黄褐色、蓝色至黑色;下皮层存在,连续。
子囊盘大多数时较多,有时很少,主要集中在地衣体中央部位,固着,直径0.5~2 mm。盘面淡黄褐色或红棕色,不被粉霜。上子实层淡褐色至黑棕色,具淡黄色颗粒,15~25 μm厚;子实层无色,50~70 μm厚;侧丝顶端膨大,无色;下子实层无色至淡灰色,20~40 μm厚;子囊棍棒状,含8孢子;子囊孢子无色,椭圆形至宽椭圆形或卵球形,8~13×4.5~6 μm。分生孢子器埋生。
化学反应:地衣体皮层K-,C-,KC+黄,P-。
化学成分:usnic acid,isousnic acid,placodiolic acid。髓层具zeorin,terpinoids。
生境:生长于各种性质的岩石上。
分布: 主要分布在干旱半干旱地区,欧洲西部,亚洲,非洲,北美西部,南美[27]。
本实验室保存的P.garovaglii标本在外部形态和子囊盘颜色、地衣体边缘裂片的凸出及波状、弯曲情况、边缘顶端的大小和宽窄等方面与其他地区报道的标本存在较多的差异,这可能是生长环境差异的缘故。在天山西部分布的种类子囊盘盘面淡红棕色至淡黄棕色或淡黄色。地衣体边缘具很明显的折成扇状、有褶的(具沟)、波状,变长。
引证标本:新疆乌鲁木齐县:南山小渠子林场,20100711,2088,43°28′08″N,87°54′87″E,热衣木·马木提,20201540,20107237;阿吾赞沟,20070730,1470,44°08′22″N,81°42′35″E,热衣木·马木提,20071721,20120775,20120740;伊犁:巩乃斯林场,20180817,2640,43°32′12″N,84°23′25″E,热衣木·马木提,20181615-B,20181596-A;新疆乌鲁木齐县:八一林场,20090410,1304,43°35′00″N,87°05′00″E,热衣木·马木提,20201542;一号冰川,20201014,2370,43°10′40″N,87°07′15″E,热衣木·马木提,YH0045,YH0024,YH0070,20201543;新疆乌鲁木齐:西山农场,20101020,983,43°47′52″N,87°28′11″E,热衣木·马木提,20201541,20201544;阿图什乌恰县:玉其塔什草原,20160805,3200,40°13′57″N,74°41′36″E,热衣木·马木提,201617500,201617217,201617287,201617197,201617244,201617265,201617155,201617166,201617221,201617229,201617234。
2.2.3 石墙原类梅Protoparmeliopsismuralis(Schreb.) M. Choisy, Contr. Lichénogr.1:tab.1(1929) (图版Ⅱ,H1-H4)
该种在研究区域内广泛分布。地衣体裂片状,常几个聚集在一起生长形成种群,近整齐圆形莲座状,连生至及其周围也能见到圆形或不规则、大小不等的鳞片,一般贴生,中部鳞状龟裂,边缘放射状排列。上表面颜色多样,呈淡黄色、淡黄绿色或淡绿黄色、淡褐色、淡黑绿色,有时呈灰白色或白色,很少光泽,平坦或略皱状;子囊盘多,发生在裂片边缘,常聚在地衣体中间;颜色多样,常呈黄褐色至褐色或红褐色,有些时候部分区域呈黑色。
化学反应:地衣体皮层K+淡黄色,C-,KC+黄,P-。
化学成分:usnic acid,atranorin,triterpenes,fatty acids。
生境:生长于岩石、朽木、藓土、土壤等基物上。
分布: 欧洲,亚洲,澳大利亚,印度,北美,南美[27],中国。国内分布于北京,上海,陕西,云南,福建,内蒙古,江苏,安徽,浙江,新疆。
引证标本:乌鲁木齐县:一号冰川,20201024,2370,43°10′40″N,87°07′15″E,热衣木·马木提,20201535,20201536,20201537,20201538,20201539,20201531,20201532,20201533,YH0059,YH0018,YH0065;米泉哈熊沟,20130702,1708,43°49′25″N,87°58′90″E,热衣木·马木提,20132546,20132617,20132615,20201534,20132854,20201528,20130092;和静县:巩乃斯林场,20180816,2150,43°14′47″N,84°40′55″E,热衣木·马木提,20181592,20181618,20181617,20181619;和静县:巴音布鲁克,20130729,2639,43°04′25″N,84°06′45″E,热衣木·马木提,20132386-a,20133013;乌鲁木齐县:南山小渠子林场,20100919,1880,43°29′63″N,87°65′96″E,热衣木·马木提,20201527;乌鲁木齐:石人沟,20131027,1263,43°46′18″N,87°49′72″E,热衣木·马木提,20137271;阿勒泰边防站,20140518,1075,47°35′10″N,88°39′36″E,热衣木·马木提,20140038;米泉林场,20150627,1509,43°49′71″N,87°56′86″E,热衣木·马木提,20155525;乔尔玛,20130731,3437,43°28′31″N,84°26′67″E,热衣木·马木提,20132483;伊宁县:阿吾赞沟,20070730,1450,44°08′28″N,81°42′35″E,热衣木·马木提,20077235;木垒东城,20171004,1856,43°42′25″N,90°10′94″E,热衣木·马木提,20176583;乌鲁木齐县:西白杨沟,20191006,2190,43°45′59″N,87°36′39″E,热衣木·马木提,20191701;新源那拉提,20070708,1540,43°17′24″N,84°12′54″E,热衣木·马木提,20071815。
2.2.4 盾原类梅Protoparmeliopsispeltata(Ramond),Arup, ZhaoXin, Lumbsch, FungalDiversity, 78(1):301(2015)(图版Ⅱ,J1-J4)
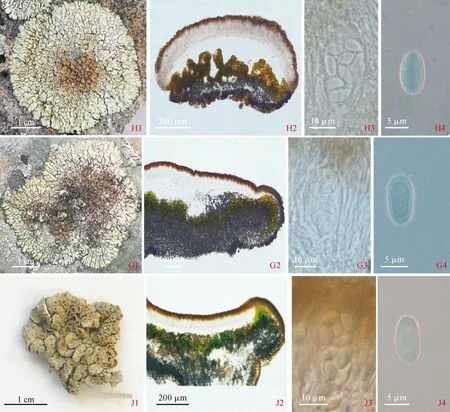
图版Ⅱ 原类梅地衣其他种类G1-G4.戛氏原类梅(标本号20181615):G1.地衣体外部形态结构;G2.子囊盘纵切解剖结构;G3.子囊;G4.子囊孢子。H1-H4.石墙原类梅(标本号20077235): H1.地衣体外部形态结构;H2.子囊盘纵切解剖结构; H3.子囊;H4.子囊孢子。 J1-J4.盾原类梅(标本号20132366):J1.地衣体外部形态结构;J2.子囊盘纵切解剖结构;J3.子囊;J4.子囊孢子Plate Ⅱ Species of Protoparmeliopsis G1-G4. Protoparmeliopsis garovaglii (Herb. No.: 20181615): G1. Morphological structure of thallus; G2. Cross section of apothecium; G3. Asci; G4. Ascospores. H1-H4. Protoparmeliopsis muralis(Herb. No.:20077235): H1. Morphological structure of thallus; H2. Cross section of apothecium; H3. Asci; H4. Ascospores. J1-J4. Protoparmeliopsis peltata(Herb. No.:20132366): J1. Morphological structure of thallus; J2. Cross section of apothecium; J3. Asci; J4.Ascospores
地衣体脐状,多叶状,3 cm宽,扁平至稍微内凹,全面至粗糙和锯齿状;上表面呈淡绿黄色至黄色,连续至龟裂,不被粉霜至部分被粉霜,末端与地衣体同色或黑色;下表面呈淡黄棕色,向末端呈淡蓝黑色,平滑至不平坦,地衣体中间常裂开。
子囊盘众多,埋生至贴生,杯状或漏斗状;盘面呈淡黄褐色至淡黄色或淡红棕色,不被粉霜;边缘始终存在,全面至弯曲或粗糙的锯齿状,与地衣体同色;子实层无色,内有淡棕色颗粒;侧丝顶端无色,棍棒状;子囊孢子椭圆型至亚球状,9~12 × 5~8 μm;分生孢子器很少或无;分生孢子丝状。
化学反应:皮层K-,C-,KC+黄,P-;髓层:K-,C-,KC-,P+褐色。
成分:usnic acid,zeorin。
生境:岩面。
分布:欧洲南部,亚洲中部,非洲南部(Ryan,2002)[28]。
P.peltata常与Rhizoplacamelanophthalma混淆;P.peltata子囊盘盘面一般不被粉霜,呈淡棕色;此外,裂片的上、下表面和杯状或漏斗状子囊盘也是鉴别该种的重要特征。
引证标本:乌鲁木齐:西山农场,20101020,996,43°47′52″N,87°28′13″E,热衣木·马木提,201008003,20108003,20108000,20108005;米泉林场,20150630,2040,43°48′77″N,87°59′84″E,热衣木·马木提,20155582,20155578;乌鲁木齐县:一号冰川,20201024,2370,43°10′40″N,87°07′15″E,热衣木·马木提,YH0071,20088000;和静县:巴音布鲁克,20130730,2597,43°08′40″N,84°20′21″E,热衣木·马木提,20132366,20138006;巩乃斯林场,20180816,2510,43°14′47″N,84°40′55″E,热衣木·马木提,20181587;和静县:乔尔玛,20130731,3445,43°28′21″N,84°26′67″E,热衣木·马木提,20201500;乌鲁木齐县:南山小渠子林场,20100709,3200,43°28′47″N,87°04′07″E,热衣木·马木提,20108019;乌鲁木齐县:白杨沟,20050618,2200,43°45′59″N,87°36′39″E,热衣木·马木提,20058000;米泉哈熊沟,20130703,2036,43°48′43″N,87°59′03″E,热衣木·马木提,20138023。
