高效降解牛奶过敏原蛋白酶菌种筛选及其重组植物过氧化氢酶的功能验证
2021-07-24堵国成
汤 恒, 张 娟, 堵国成
(江南大学 生物工程学院/工业生物技术教育部重点实验室,江苏 无锡214122)
人乳的蛋白质质量浓度与动物源乳制品有较大差异(见表1),这使得免疫系统容易将非人源的蛋白质判定为过敏原因子,并引起一系列超敏反应[1]。酪蛋白作为牛乳蛋白质的主要成分大约占总蛋白质质量分数的80%,其中许多组分并不存在于母乳之中,如αs1酪蛋白、αs2酪蛋白和γ酪蛋白,这些组分很难被人体内蛋白酶消化水解,并可能以完整的结构进入人体血液中引起过敏反应[2]。目前普遍认为αs1酪蛋白是主要的过敏原[3],从表1也可以看出,αs1酪蛋白是人乳和牛乳中差别最大的蛋白质。

表1 人乳和牛乳的成分比较Table 1 Composition of human and bovine milk
αs1酪蛋白作为过敏原极难被降解的原因在于乳制品中酪蛋白相互之间极易形成酪蛋白胶粒,一般由αs1酪蛋白、αs2酪蛋白、β酪蛋白和κ酪蛋白组成,这些多为分散的、大的、近似球形的胶体粒子,直径一般在50~600 nm之间[4]。有研究表明,酪蛋白胶粒中β酪蛋白和κ酪蛋白可形成大量分子间二硫键,而巴氏灭菌的高温可使得这些二硫键更为复杂,这使得胶粒更难被蛋白酶降解[5]。
一般破坏二硫键需要合适的碱性环境和二硫还原酶或与二硫还原酶功能相似的化学还原剂。二硫还原酶包括烷基过氧化氢还原酶[6]、硫氧还原蛋白还原酶[7]和二氢硫辛酸脱氢酶[8]等;化学还原剂包括β-巯基乙醇、巯乙酸和二硫苏糖醇等。此外在生物水解中,许多角蛋白分解菌可生成亚硫酸盐破坏角蛋白酶中的二硫键,从而辅助水解过程的进行[9]。
过氧化氢酶主要存在于细胞及组织内的过氧化体中,参与许多重要反应,是生物体维持稳态的关键酶,也被广泛应用于食品工业,如维持还原环境防止食物被氧化[10]。枯草芽孢杆菌中的植物过氧化氢酶是为数不多的胞外分泌型过氧化氢还原酶,可以催化过氧化氢分解成氧和水。David等[11]的研究表明,其主要功能为协助细胞及细胞分泌的胞外酶对抗环境中的氧化应激。本研究中结合ELISA和质谱鉴定,比对两株种属相同的野生菌发酵液酶活,以及胞外酶的差异,发现了枯草芽孢杆菌植物过氧化氢酶除已报道功能外,还具有破坏酪蛋白胶粒的二硫键,协助蛋白酶高效水解过敏原αs1酪蛋白。此外,我们还将过氧化氢酶与蛋白酶混合水解脱脂奶粉,使用HPLC和LC-MS/MS分析了水解产物的口感和所生成的功能性多肽,为未来使用混合酶制剂制备脱敏奶制品提供一定理论指导。
1 材料与方法
1.1 材料
1.1.1 菌种与载体E.coliDH5α和E.coliBL21(DE3)感受态细胞:生工生物工程(上海)股份有限公司产品;枯草芽孢杆菌S7(菌种保藏编号CCTCC NO.M2016532):中国典型培养物保藏中心;原核表达载体pET-24a(+):Novagen公司产品。
1.1.2 主要试剂 细菌小量质粒提取试剂盒、细菌基因组提取试剂盒、卡那霉素和Taq DNA聚合酶:生工生物工程 (上海)股份有限公司产品;PrimerSTAR DNA聚合酶、SYBR Premix Ex Taq Kit和DNA连接酶:大连宝生物工程有限公司产品;Human casein alpha s1(CSN1S1)ELISA试剂盒:武汉华美生物工程有限公司产品;NuPAGE Bis-Tris蛋白质预制胶和脱脂奶粉:赛默飞世尔科技(中国)有限公司产品;其余试剂均为分析纯。
1.1.3 仪器与设备 高通量全自动菌种筛选及蛋白质结晶工作站(移液工作站):瑞士帝肯公司产品;MALDI-TOF/TOF基质辅助激光解析电离串联飞行时间质谱仪:美国布鲁克·道尔顿公司产品;游离氨基酸分析专用高效液相色谱仪:美国安捷伦科技有限公司产品;Q ExactiveTMPlus混合四极轨道质谱仪:美国赛默飞世尔科技有限公司产品。
1.2 方法
1.2.1 目标菌的高通量筛选 将从奶牛牧场收集到的土样处理后,初步获得3 000株野生菌,使用高通量全自动菌种筛选工作站将野生菌转移至筛选平板(200 g/L马铃薯煮烂后经6层纱布过滤获得的滤过液,20 g/L葡萄糖,15 g/L脱脂奶粉,15 g/L琼脂粉),挑选产生水解圈的野生菌划线转移至新的平板(见图1)。

图1 脱脂奶粉平板筛选Fig.1 Skim milk plate screening
1.2.2 针对底物αs1酪蛋白选择性的ELISA分析将各野生菌编号并转移至PDA液体培养基(200 g/L马铃薯煮烂后经6层纱布过滤获得的滤过液,20 g/L葡萄糖)中,30℃恒温培养至指数生长期,取发酵液上清液测定酶活进行复筛鉴定。先通过OPA法测定针对脱脂奶粉的酶活,再逐步稀释各个野生菌发酵液上清液,使其针对脱脂奶粉的酶活达到同一水平,以减少发酵液复杂成分带来的影响,此后继续以脱脂奶粉作为底物重复相同的水解过程,但使用针对αs1酪蛋白的ELISA试剂盒检测各个发酵液上清液对于特定过敏原的酶活,以确定各个野生菌对于过敏原的选择性高低。
1.2.3 发酵液上清液蛋白质的肽指纹图谱分析将各个野生菌发酵液上清液进行SDS-PAGE分析,对不同相对分子质量大小的胶条进行预处理后,与胰蛋白酶混合,37℃酶解过夜,处理后点靶进行质谱分析,质谱结果与mascot数据库进行肽段比对鉴定。
1.2.4 重组植物过氧化氢酶的构建 以提取的枯草芽孢杆菌S7基因组为模板,根据飞行质谱结果获得的植物过氧化氢酶基因序列设计引物(见表2),PCR扩增从而获取目的基因片段。随后将载体pET-24a(+)与PCR产物分别与相对应的限制性内切酶混合,在37℃下孵育1.5 h,两者酶切产物经过胶回收纯化后用DNA连接酶连接,在16℃下连接6~8 h,连接产物通过热激法转化E.coliDH5α感受态细胞,在LB卡那霉素抗性平板上挑取阳性克隆子,菌落PCR验证后提取质粒,经过测序鉴定,最终获得含有目的基因的重组质粒pET-24a(+)/katA。将质粒pET-24a(+)/katA转化E.coliBL21(DE3)感受态细胞,最终获得重组植物过氧化氢酶表达菌株。

表2 PCR引物序列Table 2 PCR primers sequence
1.2.5 重组蛋白的纯化 纯化过程全部在4℃下进行,将离心收集的重组菌充分倒入培养基。将菌体重悬于25 mL缓冲液A(200 mmol/L Tris-HCl(pH 7.4),500 mmol/L NaCl,5 mmol/Lβ-mercaptoethanol)中,使用Branson Sonifier 450对细胞进行了超声破壁处理,高速离心后收集破碎液上清液。再采用AKTA Start(GE Healthcare)和HisTrap亲和柱对目标蛋白质进行纯化,在经缓冲液A平衡后接入上清液,先使用含5 mmol/L咪唑的缓冲液A进行清洗后,使用含300 mmol/L咪唑的缓冲液A进行洗脱,获得重组植物过氧化氢酶。
1.2.6 脱脂奶粉水解产物的LC-MS/MS活性多肽鉴定 水解产物经过超滤、脱盐等预处理后,直接加载到反相预柱(Acclaim PepMap RSLC)进行肽段分离,分离后肽段经Q ExactiveTMPlus混合四极轨道质谱仪分析,收集的串联质谱结果上传至mascot搜索引擎进行分析,比对后获得对应的肽段序列。
2 结果与分析
2.1 针对底物αs1酪蛋白高选择性的野生菌筛选与鉴定
将筛选获得的野生菌进行16S rRNA和功能基因gyrA鉴定,以确定各个野生菌的种属,再分别进行发酵培养,获得其发酵液上清液。由于不同野生菌蛋白酶表达量不同,无法直接使用发酵液上清液进行横向比较。而OPA法可检测整个体系水解程度,ELISA可以检测水解反应体系中针对某一特定底物的水解程度,利用此特点设计实验,首先通过OPA法确定各个发酵液的酶活,再分别稀释,使其针对整体底物脱脂奶粉的酶活达到相同水平。此后,将调整浓度后的发酵液对脱脂奶粉进行水解反应,并通过ELISA分析对底物αs1酪蛋白的酶活。如此可筛选出对混合底物脱脂奶粉的酶活相同时,对过敏原αs1酪蛋白选择性更高的蛋白酶产生菌。如图2所示,横坐标为各个野生菌的种属名和筛选编号,纵坐标为调整后发酵液上清液的水解αs1酪蛋白酶活。其中酶活最高的为枯草芽孢杆菌S7(Bacillus subtilisS7)发酵液上清液,而作为对照的商业酶A-2SD和NY-10酶活较低。在总酶活调整一致的情况下,其他野生菌针对过敏原酶活水平不高,如嗜麦芽寡养单胞菌S2、绿脓假单胞菌S8(非食品安全菌),故选择枯草芽孢杆菌S7为去除过敏原的主要研究对象,并进行菌种保藏(CCTCC NO.M2016532)。同时被鉴定为枯草芽孢杆菌的还有S21和S23,值得关注的是S21和S23酶活水平接近,但与S7有显著差异。
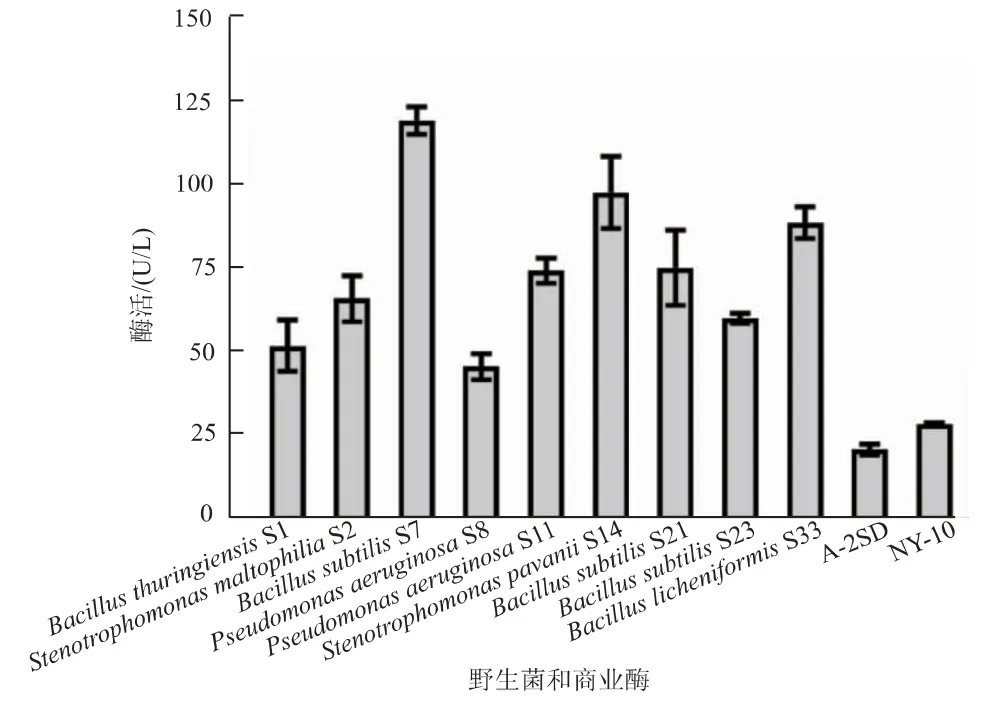
图2 针对αs1酪蛋白作为底物的酶活性比较Fig.2 Activities of each enzyme usingαs1-casein as the substrate
2.2 关键酶鉴定
通过比对S7和S21的发酵液上清液进行SDS-PAGE分析发现,枯草芽孢杆菌S7在5.5×104位置分泌的一种蛋白质在S21发酵液中分泌量极少。此后使用MALDI-TOF/TOF对各个条带进行肽指纹图谱分析,如图3所示,以3.2×104的鞭毛蛋白(UniProtKB:P02968,含量最高条带)作为对比参照,主要蛋白酶为2.8×104的枯草芽孢杆菌蛋白酶E(UniProtKB:P04189),而表达量差异最大是5.5×104的植物过氧化氢酶(UniProtKB:P26901)。

图3 S7和S21发酵液上清液SDS-PAGE分析Fig.3 SDS-PAGE analysis of the supernatants after S7 and S21 fermentation
2.3 植物过氧化氢酶的重组表达及功能验证
通过从枯草芽孢杆菌S7中调取植物过氧化氢酶基因,并构建重组表达质粒,获得异源表达菌株。再通过诱导表达和镍柱亲和层析法,获得了重组植物过氧化氢酶。如图4中SDS-PAGE所示,泳道1为镍柱纯化流穿峰,泳道2为5 mmol/L咪唑洗脱峰,泳道3为300 mmol/L咪唑洗脱峰,在5.5×104左右可见明显条带,证明获得可融性表达的重组酶。

图4 重组枯草芽孢杆菌S7植物过氧化氢酶的镍柱纯化SDS-PAGE分析Fig.4 SDS-PAGE analysis of recombinant vegetative catalase from Bacillus subtilis S7 nickel column purification
通过将纯化后的重组植物过氧化氢酶回补至S21发酵液上清液中,S21水解αs1酪蛋白的酶活从75.43 U/L提升至106.18 U/L达到与S7的119.43 U/L相近水平,从而证明了S7和S21的酶活差异主要是由植物过氧化氢酶的表达量差异导致的。据此,我们发现植物过氧化氢酶不仅可以水解过氧化氢,维持更好的还原环境,协助枯草芽孢杆菌蛋白酶E抵抗氧化应激,保护其结构不被自由基破坏,增加蛋白酶作用时间;可能还具有破坏蛋白胶粒中的二硫键,将其中的过敏原αs1酪蛋白释放出来,使蛋白酶可以更高效地进行水解反应,增强蛋白酶的选择性脱敏效果(见图5)。
2.4 植物过氧化氢酶对风味的影响
采用已报道的高效液相色谱法[12]测定S7和S21发酵液上清液水解脱脂奶粉后水解产物中游离氨基酸的浓度,并比对二者差别。如图6所示,两种枯草芽孢杆菌的发酵液上清液可水解脱脂奶粉生成氨基酸比例相似的水解产物,如均可生成较多亲水氨基酸,如谷氨酸(E)和酪氨酸(Y),以及较多的疏水氨基酸,如缬氨酸(V)、亮氨酸(L)和苯丙氨酸(F)。但由于S7发酵液上清液中含有较多的植物过氧化氢酶,可打开二硫键使得更多的水解位点得以暴露,有效增加蛋白酶对酪蛋白胶粒,特别是其中αs1酪蛋白的水解,同时也就形成了对风味更为有利的水解产物,如相比于S21生成了较多亲水氨基酸赖氨酸(K),且生成的疏水氨基酸浓度和比例也呈显著下降。
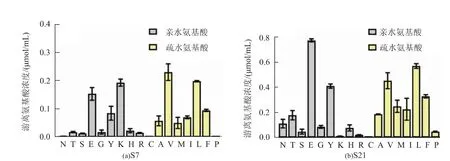
图6 S7和S21发酵液上清液水解脱脂奶粉后水解产物中游离氨基酸对比Fig.6 Comparison of free amino acids in the skim milk hydrolysate between S7 and S21
水解产物中的苦涩风味主要来自各类疏水氨基酸,如亮氨酸(L)和苯丙氨酸(F),而甲硫氨酸(M)则会带来类似肉类、蛋类的不良风味[13],这对于奶粉的制备是不利影响。为验证其风味,邀请5位品鉴人对微量S7和S21水解5 h后的脱脂奶粉水解液进行感官评定实验,并给予0~5分的打分。如表3所示,相比于未处理的脱脂奶粉,两种水解液苦味均有增加,但S7的水解液苦味较低更易被接受。由于植物过氧化氢酶的加入,水解产物中的亲水氨基酸增多且疏水氨基酸减少,这使得水解后的脱敏奶制品具有更易接受的风味及应用价值。
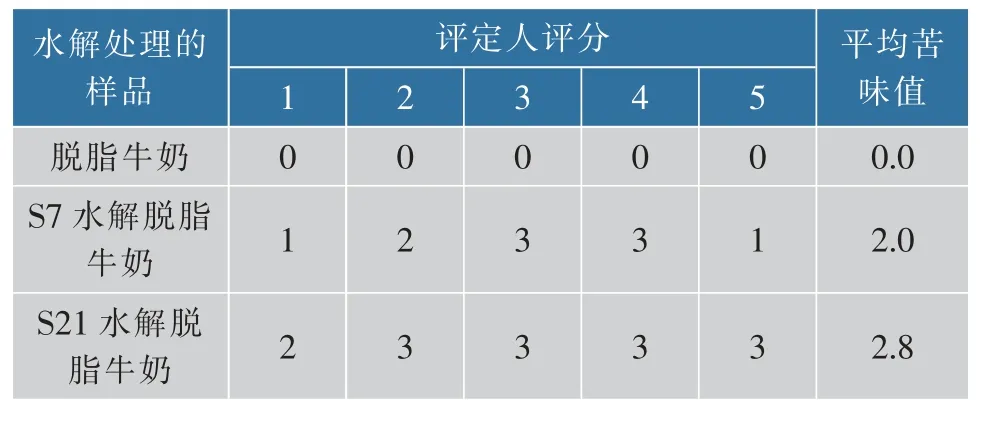
表3 感官评定Table 3 Sensory evaluation
2.5 脱脂奶粉水解产物的功能性多肽鉴定
将枯草芽孢杆菌S7的脱脂奶粉水解产物进行LC-MS/MS活性多肽鉴定,在获得肽段序列后与已报道的文献进行比对。如表4所示,水解产物中含有抗高血压活性多肽5种,以及抗菌活性多肽、矿物质结合活性多肽、血管紧张素转化酶抑制活性多肽、免疫调制活性多肽和抗癌活性多肽各1种,这些肽在天然蛋白质中并无活性,在被水解释放后才具有活性。他们可以以完整的形式被吸收,在肠道局部发挥各种生理作用,也可以在进入循环系统后产生全身作用[14]。进一步证明植物过氧化氢酶和枯草芽孢杆菌蛋白酶E混合水解的脱敏奶制品具有更好应用价值。

表4 功能性多肽鉴定Table 4 Identification of functional peptides
3 结语
αs1酪蛋白是奶制品的主要过敏原,人体自身消化系统难以水解,从而导致该过敏原可完整的通过消化系统进入血液从而诱发人体过敏反应。αs1酪蛋白难以被水解的主要原因在于各种酪蛋白容易通过疏水作用和分子间二硫键相互交联形成酪蛋白胶粒,使得过敏原αs1酪蛋白被包裹其中,蛋白酶难以接触其水解位点。化学还原剂可有效破坏各类二硫键,但难以应用于食品工业中。相比而言,生物还原剂则具有用量少、可高温灭活去除等优点,如烷基过氧化氢还原酶、硫氧还原蛋白还原酶和二氢硫辛酸脱氢酶等均被报道可有效破坏二硫键,但这些二硫还原酶均为胞内酶难以适应胞外复杂环境;Sri等[22]报道了来自Bacillussp.MTS的一种分泌型二硫还原酶,但其原始菌非食品安全菌。此外,使用其他来源的分泌型二硫还原酶极可能与筛选获得的枯草芽孢杆菌蛋白酶E不适配,导致其被蛋白酶水解而无法发挥作用,故挑选枯草芽孢杆菌植物过氧化氢酶辅助水解。在此前的报道中,此酶已被多次证实可帮助细胞和分泌酶对抗氧化应激[11,22],本实验结果表明在其存在的情况下,枯草芽孢杆菌蛋白酶E对于水解αs1酪蛋白的酶活可增强约1.5倍,从而推测该酶可促进蛋白酶水解富含二硫键的底物。将获得的枯草芽孢杆菌蛋白酶E和枯草芽孢杆菌植物过氧化氢酶混合,应用于脱敏奶制品制备,其产品除了具有低过敏原,还有相对容易接受的口感并含有一定的生物功能性多肽,具有较好的食品工业应用价值。
