鲤Cyprinus carpio、泥鳅Misgurnus anguillicaudatus与黄豆种子共生系统对水质的影响
2021-07-24刘帅袁胜煜魏海峰刘长发
刘帅,袁胜煜,魏海峰,刘长发
(1.辽宁省近岸海洋环境科学与技术重点实验室,辽宁 大连 116023;2.大连海洋大学海洋科技与环境学院,辽宁 大连 116023;3.大连海洋大学农业农村部北方海水增养殖重点实验室,辽宁 大连 116023)
鱼菜共生系统是一种新型养殖系统,利用鱼类和蔬菜协同共生的生态关系,开发了水产养殖业和无土栽培业相结合的综合性养殖生态工程技术[1-3]。鱼菜共生系统中,植物对水体具有一定的吸收净化作用,可改变水体的理化指标,使其达到鱼类生长的适宜条件,促进鱼类生长。相关研究证明,鱼菜共生系统中的植物可将水体中85%左右的悬浮物质和75%左右的有机物质除去,降低水生物耗氧量达90%以上[4],鱼菜共生养殖系统已被许多国家深度推广[5,6]。黄豆为水培植物,生长速度快于其他植物,短时间即能获取更大的收益;黄豆价格低廉、取材较易,是非常合适的实验对象。目前,利用黄豆作为水培植物的相关报道空缺,故本文将黄豆作为创新性鱼菜共生系统中的水培植物,这为鱼菜共生系统中水培植物的选择提供了基本理论和科学线索。
研究初期,主要研究了鱼菜共生系统中植物密度[7]、植物种类[8]、饲料铁元素含量[9]以及每日喂食量比例[10]对鱼类和植物的影响,但对鱼菜共生系统中水质的分析研究尚未完善。水是水生生物生存的介质,特别是生活在水中的鱼类,它们从出生到死亡整个生命活动过程都在水体中度过。因此,研究鱼菜共生系统中水质的变化规律,调控水体理化因子,有利于提高鱼类的成活率,加速养殖业的发展[11]。本文通过研究鱼菜共生系统对水体理化性质的影响,可为进一步了解鱼菜共生系统作用机制、完善科学理论、改进鱼菜共生系统提供基本理论和科学依据。
1 材料与方法
1.1 材料
试验用健康鲤Cyprinus carpio 和泥鳅Misgurnus anguillicaudatus,初始质量分别为(1.5±0.2)kg和(70±2)g。黄豆种子购置大连黑石礁农贸市场。每个黄豆育苗盘30 cm×40 cm,内放黄豆50 g,漂浮在玻璃槽中,每层三个育苗盘。给试验鱼投喂饲料,每天喂食2 次,日投喂量为鱼体质量的2%,摄食结束后捞出残余饲料。
实验室中构建的鱼菜共生系统由三层结构组成:第一层为137 cm×60 cm×50.7 cm 鱼缸,体积约为277.8 L,用于饲养鱼;第二、三层为134.5 cm×51 cm×8 cm 的玻璃槽,用于种植水培植物,三层顶部都装有LED 灯(红蓝比为8∶2),每天光暗比为12∶12。用90 W 功率水泵将鱼缸内的养殖水体提升至第二、三层玻璃槽,经植物根部过滤后在重力作用下,经过鱼缸上过滤棉过滤后回流至鱼缸,构成完整的养殖循环系统。鱼菜共生系统循环水流在栽培槽中停留20 min。该系统水循环率达20 次/d。鱼菜生系统不与外界环境进行水体交换。
1.2 方法
试验期间共监测8 个指标:pH、温度、溶解氧、氨氮、亚硝态氮、硝态氮、总氮和活性磷含量。pH、温度由HANNA 的水温、pH 一体化测试仪(水温量程0.0~60.0℃,精度±0.5℃;pH 量程0.00~14.00,精度±0.05)测量;溶解氧含量由雷磁溶氧仪(量程0.00~20.00 mg/L,精度±1.5%)测量;氨氮浓度由纳氏试剂分光光度法测定,亚硝态氮含量由盐酸萘乙二胺分光光度法测定,硝态氮含量由紫外分光光度法测定,总氮含量和活性磷含量分别按照HJ636-2012《水质总氮的测定碱性过硫酸钾消解紫外分光光度法》、GB11893-1989《水质总磷的测定钼酸铵分光光度法》测定。
1.3 数据统计与处理
用Excel 软件初步整理测定的数据,绘制出相应的折线图,用SPSS 22.0 统计软件对实验数据进行统计学分析和差异性分析,P<0.05 时,表示差异显著。
2 结果与分析
2.1 鱼菜共生系统中pH 的变化规律
由图1 可知,在鱼菜共生系统中,随着时间的增加,试验水体pH 先升高(第1~7 d)后降低(第7~10 d),再升高(第10~21 d)并趋于平稳。试验水体pH 变动在7.5~7.82 之间,但最终趋于7.7 附近,较初始状态增长2.25%。在鱼菜共生系统中,鱼缸上层的过滤棉产生硝化细菌,其硝化作用会升高pH,以及水培植物的光合作用消耗二氧化碳升高pH,调节水体的酸碱度,水体pH 在7.5~7.8 之间正常波动。
2.2 鱼菜共生系统中溶解氧含量的变化规律
由图1 可知,在鱼菜共生系统中,随着时间的增加,试验水体溶解氧含量在7.0 mg·L-1左右波动,第7 d 达到峰值7.4 mg·L-1,较第1 d 增长5.71%。之后溶解氧含量上下波动,在第13 d 达到最小值6.3 mg·L-1。在13~32 d 整体呈上升状态,第32 d 达最大值7.5 mg·L-1,较第1 d 增长7.14%。这是由于鱼菜共生系统循环水回流过程中与大气中的氧气接触,增加养殖水体中氧气含量,又鱼类呼吸作用消耗水中溶氧量存在差异,故在第7~21 d 范围内,溶解氧在6.2~7.4 mg/L 范围内正常波动,可见鱼菜共生系统对水体溶解氧含量具有一定的调节作用。
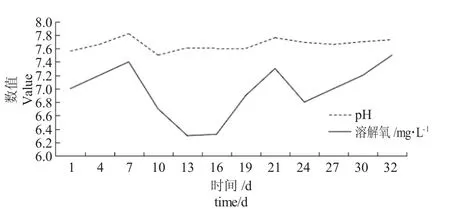
图1 鱼菜共生系统中pH 和溶解氧含量的变化规律Fig.1 Changes in pH and dissolved oxygen content in the fish and vegetable symbiotic system
2.3 鱼菜共生系统中氨氮浓度的变化规律
由图2 可知,在鱼菜共生系统中,随着时间的增加,试验水体氨氮浓度显著降低,最后处于较低的平稳值0.1136 mg·L-1,较第1 d 下降85.64%。第1~21 d 氨氮浓度与时间之间存在显著的线性相关关系,相关方程分别为:y =-0.0962x+0.8554,R2=0.9876。在鱼菜共生系统中,鱼缸上层的过滤棉产生硝化细菌,其硝化作用将氨氮转化成了亚硝酸盐,进一步转化为硝酸盐,同时水培植物也会吸收水体中的氨氮,但硝化作用是氨氮浓度降低的主要作用机制[12]。
2.4 鱼菜共生系统中亚硝态氮含量的变化规律
由图2 可知,在鱼菜共生系统中,随着时间的增加,试验水体亚硝态氮含量显著降低,最后达到最低值0.078 mg·L-1,此值较第1 d 下降82.63%。第1~32 d 亚硝态氮含量与时间之间存在显著的线性相关关系,相关方程为:y =-0.0369x+0.504,R2=0.9461。鱼类的粪便和残饵含有大量的亚硝态氮,导致鱼菜共生系统亚硝态氮含量初始处于较高状态,但系统中的水培植物可吸收亚硝态氮,水体中亚硝态氮含量逐渐降低。
2.5 鱼菜共生系统中总氮含量的变化规律
由图2 可知,在鱼菜共生系统中,随着时间的增加,鱼菜共生水体总氮含量显著降低,最后达到最低值0.0672 mg·L-1,较第1 d 下降93.32%。第1~32 d 总氮含量与时间之间存在显著的线性相关关系,相关方程分别为:y =-0.0879x+0.9882,R2=0.9507。在鱼菜共生系统中,鱼类的粪便和残饵含有大量的有机氮和无机氮,水体中的无机氮可被水培植物吸收用于生长,水体中的有机氮在亚硝化细菌的作用下发生氨化作用转化为氨氮,因此水体中的总氮含量逐渐减少,但每天都会产生的鱼类粪便和残饵,抑制了总氮含量降低的速率且最终达到平衡状态。
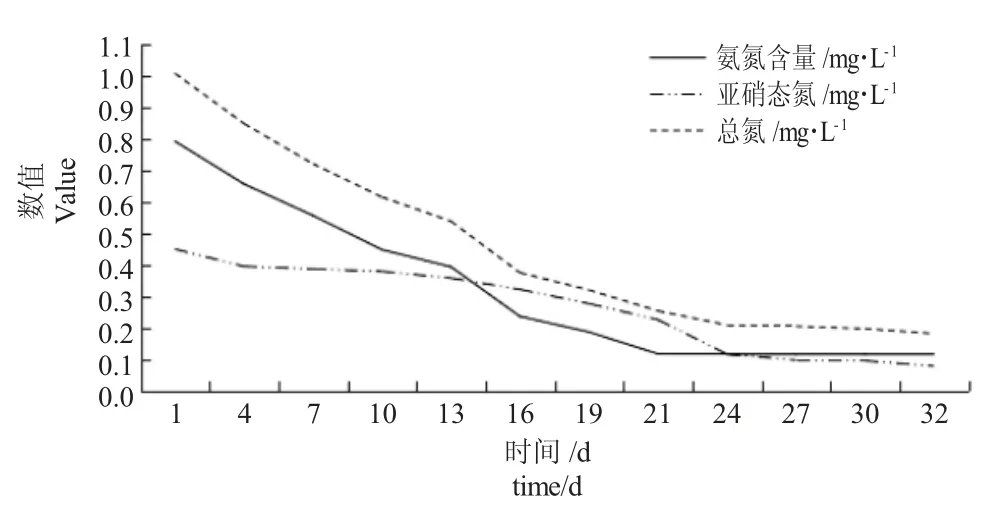
图2 鱼菜共生系统中氨氮、亚硝态氮和总氮含量的变化规律Fig.2 Changes in concentrations of ammonia nitrogen,and nitrite nitrogen content and total nitrogen content in the fish and vegetable symbiotic system
2.6 鱼菜共生系统中活性磷含量的变化规律
由图3 可知,在鱼菜共生系统中,随着时间的增加,试验水体活性磷含量显著降低,最后达到最低值1.422 mg·L-1,较第1 d 下降71.01%。第1~32 d总活性磷含量与时间之间存在显著的线性相关关系,相关方程分别为:y =-0.3226x+4.9606,R2=0.9803。在鱼菜共生系统中,活性磷主要来自鱼类的粪便和残饵,水培植物能吸收水体中活性磷用于生长发育[13],故活性磷含量逐渐降低。但随着时间的推移,养殖水中的微生物数量逐渐增多,会导致活性磷含量的增加,最终使活性磷含量达到一种相对稳定的动态平衡。

图3 鱼菜共生系统中活性磷含量的变化规律Fig.3 Changes in active phosphorus content in the fish and vegetable symbiotic system
2.7 鱼菜共生系统中硝态氮含量的变化规律
由图4 可知,在鱼菜共生系统中,随着时间的增加,试验水体硝态氮含量在第1~13 d 迅速下降,第13 d 达到最小值0.0337 mg·L-1,较第1 d 下降55.36%;第13~21 d,硝态氮含量显著上升,第21 d达到峰值0.0666 mg·L-1,较第1 d 下降11.79%;第21~32 d,硝态氮含量再次显著下降,第32 d 达到谷值0.0666 mg·L-1,较第1 d 下降42.65%。在鱼菜共生系统中,鱼类的粪便和残饵含有大量的硝态氮,水培植物可吸收水体中硝态氮用于自身发育生长,故出现水体硝态氮含量在0.0337~0.0755 mg·L-1正常波动的现象。
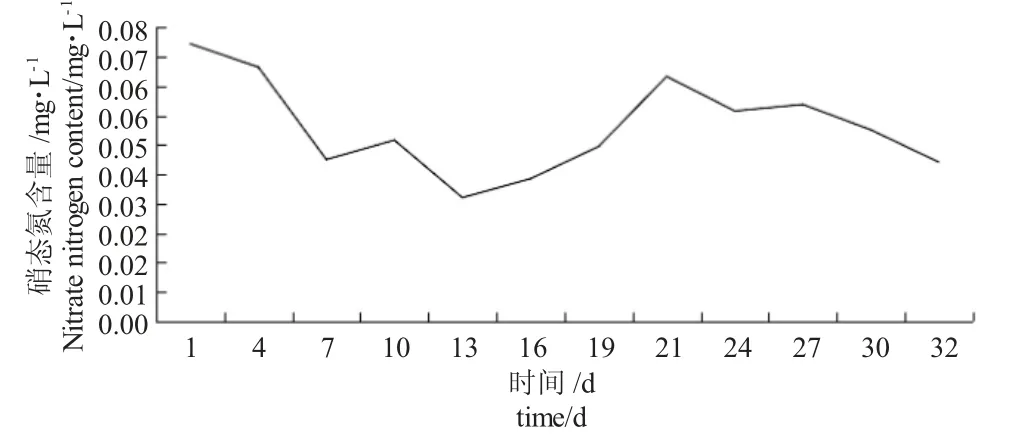
图4 鱼菜共生系统中硝态氮含量的变化规律Fig.4 Changes in nitrate nitrogen content in the fish and vegetable symbiotic system
3 讨论
实验水体水质指标检测结果的显著性分析表明,鱼菜共生明显改良了水质,pH 略微升高但保持在适宜养殖的范围内,水温上升3 ℃左右,溶解氧含量增长0.5 mg·L-1,氨氮、亚硝态氮、硝态氮、总氮和活性磷浓度分别降低了85.64%、82.63%、42.65%、93.32%和71.01%。
不同生物的最适生长pH 范围不同[14],维持养殖水pH 稳定对促进鱼菜共生系统中各生物组分的生长具有重要意义。合适的pH 范围能滋生硝化细菌,其硝化作用将对鱼类有毒性作用的氨氮、亚硝氮转化成低毒性的硝态氮,提高了鱼类的存活率[15]。李晓辉等[16]报道,池塘鱼菜共生系统可改善水质,使水体pH 稳定在7.5~8.0 之间,达到鱼类和植物适宜的环境。蔡淑芳等[17]研究显示,池塘鱼菜共生系统可使水体pH 稳定在7.5~8.0 之间,大幅改善了鱼类和植物的生长。张文良等[8]证明,当池塘鱼菜共生系统维持池塘水体pH 为7.0~8.0 之间时,鲫Carassius auratus 苗的存活率最高。本实验中,水体pH 出现先升高(第1~7 d)后降低(第7~10 d),再升高(第10~21 d)并趋于平稳的状态。水体pH 在7.5~7.8 之间变动,一直维持在适宜鱼类养殖的范围内。
水中溶解氧是水生动物赖以生存的必要条件之一,大部分水域溶解氧含量在8~10 mg·L-1,低氧会导致鱼类缺氧甚至窒息死亡。水中溶解氧含量也是限制反硝化和硝化过程的主要因素之一,低溶解氧含量不能将具有毒性的亚硝酸盐氮(NO2-N)氧化,极大影响反硝化作用所需酶的合成和表达以及N2O 还原酶的活性[18]。施吉华[11]发现,水体溶解氧含量高能提高鱼类的摄食量,促进鱼类生长。本研究中,实验水体溶解氧含量在第1~7 d 缓慢上升,之后上下波动,在第13~32 d 整体出现上升的状态,第32 d 出现最大值7.5 mg·L-1,较无鱼菜共生系统增长7.14%,可见鱼菜共生系统可提高水体溶解氧含量。
在池塘养殖中,较高的氨氮和亚硝态氮含量会抑制鱼类的生长,甚至导致鱼类大面积死亡[19]。李波等[20]研究了不同溶氧水平下氨氮和亚硝酸盐对黄颡鱼Pelteobagrus fulvidraco 的急性毒性,发现较高的氨氮浓度和亚硝态氮含量损坏水生动物的鳃组织,抑制呼吸作用和呼吸速率,破坏了生物体内外气体交换,导致生物中毒。运珞珈等[21]研究发现,较高的氨氮和亚硝态氮含量会导致稚鳖Trionyx sinensis 养殖水体中病菌大量繁殖,污染了养殖水体,使养殖的鳖致死。本实验中,水体氨氮浓度和亚硝态氮含量显著降低,分别达到最低值0.1136 mg·L-1和0.078 mg·L-1,远远低于起始的氨氮浓度和亚硝态氮含量。
在鱼类生长的水环境中,活性磷含量不是主要的影响因素,但活性磷含量会引起养殖水体富营养化,抑制鱼类的生长发育[22]。本实验初期,鱼菜系统实验水体活性磷含量为4.9 mg·L-1,随时间的增长,水体活性磷含量呈明显梯度下降,最终达到最低值1.42 mg·L-1。这与闫晓君[23]的鱼菜共生系统中微生物的作用使活性磷含量逐渐下降现象一致。
鱼菜共生系统合理地控制了养殖水体pH、增加溶解氧含量、大幅度降低氨氮、亚硝态氮、硝态氮、总氮、活性磷含量,较大程度上为鱼类提供适宜发育环境,是养殖业的一大突破。水培植物的水质净化、修复等功能,使总氮含量和磷酸盐处于低水平稳定状态,鱼菜共生系统可以长时间不换水,不排污,实现污染物零排放,将是环保行业的一大契机。
