蓝点马鲛渔业生物学研究进展
2021-07-21瞿俊跃杨光明媚陈新军
瞿俊跃,杨光明媚,方 舟,2,3,4,5,陈新军,2,3,4,5
( 1.上海海洋大学 海洋科学学院,上海 201306; 2.大洋渔业资源可持续开发教育部重点实验室,上海 201306; 3.国家远洋渔业工程技术研究中心,上海 201306;4.农业农村部大洋渔业开发重点实验室,上海 201306; 5.农业农村部大洋渔业资源环境科学观测实验站,上海 201306 )
蓝点马鲛(Scomberomorusniphonius)俗称鲅鱼、马鲛等,属鲈形目、鲭科、马鲛属,系暖温性中上层洄游性经济鱼类[1-2],广泛分布于西北太平洋的日本海、日本太平洋沿岸、朝鲜半岛沿岸及我国的渤海、黄海、东海海域[3-6]。蓝点马鲛是我国高产的重要经济鱼种之一[7],也是日本、韩国及中国台湾等多个国家和地区的重要捕捞对象[8],中国沿海的作业方式以拖网和刺网为主,其中流刺网作业时间集中在春汛[9-10]。据日本西海区水产研究所提供的蓝点马鲛资源评估报告[11](图1),20世纪80年代以来,蓝点马鲛资源获得大规模开发利用,1993年中国、日本、韩国的蓝点马鲛总产量为1.63×105t,随后呈现迅速上升的趋势,1999年达到历史最高产量5.89×105t,之后虽有回落,但年产量仍保持在4.0×105~5.4×105t,2018年总产量为4.0×105t。在当今渔业资源严重衰退情况下[12-14],蓝点马鲛是目前我国少数维持高产的渔获种类之一[15],因此研究蓝点马鲛渔业生物学和渔业海洋学,对合理开发和利用该鱼种资源状况及科学管理具有重要意义[16]。为此,笔者结合国内外学者对蓝点马鲛的群体划分[3,17-19]、洄游习性[20-21]、年龄与生长[22-25]、摄食生态[26-30]、繁殖习性[31-33]、渔场分布[9,34-36]、资源评估管理[37-40]等方面的研究现状做了归纳总结,并分析讨论蓝点马鲛渔业生物学研究过程中存在的问题,探索今后应该努力的研究方向,以期为合理地开发和利用该资源提供依据。

图1 1984—2015年中国、日本、韩国蓝点马鲛产量[11]Fig.1 Annual catch of Japanese Spanish mackerel S. niphonius in China, Japan and South Korea from 1984 to 2015[11]
1 渔业生产情况
蓝点马鲛的产量在过去的30多年里经历了很大的变化。据日本西海区水产研究所统计[11],蓝点马鲛是中国、日本及韩国的重要捕捞对象。中国年捕捞量约占3国总量的80%以上,2000年后中国年捕捞量是日本的30余倍。1980年韩国捕捞量与日本捕捞量相同,但2000年以后,韩国捕捞量超过了日本。20世纪80年代以来,蓝点马鲛资源得到大规模开发利用,1984—1992年,中日韩3国的总捕捞量基本保持在2.0×105t,1993年中日韩3国的蓝点马鲛总产量为1.63×105t,随后呈现迅速上升的趋势,捕捞量保持在每年8×104t的增长,1999年达到历史最高产量5.89×105t,与水温上升密切相关,之后虽有回落,但年产量也均保持在4.0×105~5.4×105t,2015年总产量为4.8×105t。1992—1997年间,韩国蓝点马鲛的捕捞量较低,但1998年有所增加,此后捕捞量基本稳定在2×104~3×104t,2007年创下42 199 t的历史最高记录,之后有所下降,2016年为35 869 t。
2 种群组成与群系划分
种群是生态系统中生物群落的基本组成单元,忽视潜在的空间种群结构会增加局部种群灭绝的风险,因而鱼类管理单元的正确划分是该渔业资源可持续利用的先决条件[41-43]。关于蓝点马鲛种群问题,从20世纪80年代至今一直存在争议,笔者整理50多年来不同种群研究方法对渤海、黄海、东海蓝点马鲛种群划分的结论(表1),提出蓝点马鲛种群划分分歧所在。
韦晟等[44]主要依据腹脊椎骨数、尾脊椎骨数、背鳍鳍条数、臀鳍鳍条数和左右鳃耙数等计数性状,将渤海、黄海的蓝点马鲛分成了渤海—黄海中北部群系和黄海南部种群。黄克勤等[17]用聚类分析综合分析了计数性状和量度性状,发现东海蓝点马鲛仅有1个种群。王英俊等[18]利用传统形态分析方法测量了耳石质量及外部形态指标并进行聚类分析,结果表明,黄海的吕泗渔场、赣榆渔场及胶南渔场的蓝点马鲛均为长江口越冬场进入黄渤海区的种群。堀川博史等[45-46]根据渔业调查结果,认为渤海、黄海、东海海域存在2个种群,分别为黄海及渤海种群、东海种群。水柏年等[47]综合分析了黄海及东海5个蓝点马鲛渔场群体的21个外部形态指标,指出黄海和东海蓝点马鲛为不同的种群。Zhang等[19]分别利用耳石外部形态的传统方法和傅里叶函数法对渤海及黄海3个产卵场的蓝点马鲛种群进行了判别分析,研究表明,傅里叶形态分析法相较于传统判别有着更高的准确率,并将其分成了渤海种群和黄海中南部种群。近些年相关学者利用分子生物学方法对蓝点马鲛黄海及东海种群进行研究,线粒体DNA和CO Ⅰ序列分析认为东海、黄海种群之间遗传距离较短,不同地理种群的个体混杂分布,不存在明显地理结构,并没有明显的种群遗传分化[8,16];而核基因组多样性数据(AFLP)分析显示,东海、黄海种群又有着显著的地理种群分化[48]。
综上,蓝点马鲛种群划分主要推论可归纳为:(1)黄海及渤海种群、东海种群;(2)东海、黄海、渤海均为同一种群,但可能存在亚种群[49]。

表1 渤海、黄海、东海蓝点马鲛种群划分
3 洄游分布
蓝点马鲛为暖水性中上层鱼类,游泳能力强,活动范围大,产卵和越冬时进行长距离洄游[50-51]。其洄游研究主要利用标志放流[52]、渔业调查[21]、耳石微化学[53]等来实现。
蓝点马鲛的越冬场,主要分布于东海外海和黄海南部的济州岛附近[35],每年1—2月,蓝点马鲛在这两个海域水深80~100 m越冬,3月时各产卵群体随海流离开各自的越冬场陆续向近岸生殖洄游,东海外海越冬场群体沿中国台湾暖流北上,大部分进入长江口以南的舟山及象山港水域进行产卵,其中性成熟较早的个体,进入吕泗渔场,产卵期为4月中下旬及5月上旬;另一部分穿过索饵场继续北上,沿海岸分赴渤海和黄海中北部产卵场进行产卵洄游,产卵期为5月上旬至5月底。
济州岛越冬场群体对该水域的影响也至关重要,该群体相对于东海越冬场群体离开越冬场时间较晚,沿黄海暖流进入黄海南部的大沙渔场,部分性成熟较早个体,于4月中下旬抵黄海北部吕泗渔场产卵,主支则继续向西北洄游,进入海州湾或游向黄海、渤海其他产卵场[45],另一分支则沿着对马暖流分支进入纪伊水道、丰后水道及濑户内海产卵,产卵时间为5—7月[54]。8—9月产卵后群体和当年生仔鱼、幼鱼开始按原路返回,在索饵场分散索饵;10月左右,随着水温下降,渤海、黄海种群将南下,12月到达东海北、中部越冬场[30](图2)。

图2 我国沿岸蓝点马鲛洄游路径(改编自文献[19])Fig.2 The migration path of Japanese Spanish mackerel S. niphonius in the coast of China (revised from document[19])a.黑潮; b.对马暖流; c.黄海暖流; d.渤海环流; e.台湾暖流; f.苏北沿岸流; h.长江口冲淡水; i.东南沿岸流a.Kuroshio current; b.Tsushima current; c.Yellow Sea Warm current; d.Bohai Sea circulation; e.Taiwan warm current; f.coastal current in north Jiangsu province; h.Yangtze Diluted water; i.Southeast coastal current
4 年龄与生长
4.1 耳石特征及年龄鉴定
鲭科鱼类常用鳞片、耳石、脊椎骨和鳃盖骨作为年龄鉴定的材料[55]。但由于蓝点马鲛体表的圆鳞具有易脱落的特性,拖网作业对其影响较大[56],因而常用耳石进行年龄鉴定。耳石是一种主要由碳酸钙等组成的矿化组织,其形态比骨骼更为稳定[57]。已有学者利用蓝点马鲛耳石进行种类鉴别[18]、年龄鉴定[58]及形态生长[59]等领域的研究。
蓝点马鲛耳石近似长方形,正面内凹,背面外凸,并有一凹槽,将矢耳石分成背区、前区、后区和腹区4部分(图3)。有学者分别对东海外海海域、渤海黄海海域及象山港海域的蓝点马鲛的左、右矢耳石差异进行了分析,结果表明,蓝点马鲛左、右矢耳石各部分间无显著差异[18,60-61],因此左、右耳石均可作为蓝点马鲛耳石方面的研究材料。通过对东海外海蓝点马鲛不同叉长组耳石外部形态的研究,发现耳石形态可能受淋巴液流动和游泳速度变化的影响[62],随着个体的增长,蓝点马鲛耳石总体上呈现纵向缩小、横向增长的趋势;且研究结果显示,蓝点马鲛初次性成熟叉长可能为350~400 mm。象山港蓝点马鲛不同年龄组的耳石形态存在显著差异[60],即随着蓝点马鲛的生长,其耳石形态会发生显著变化。渤海、黄海及东海海域蓝点马鲛雌、雄个体之间耳石形态的差异未达到雌、雄二型水平[23,60],不存在显著差异。
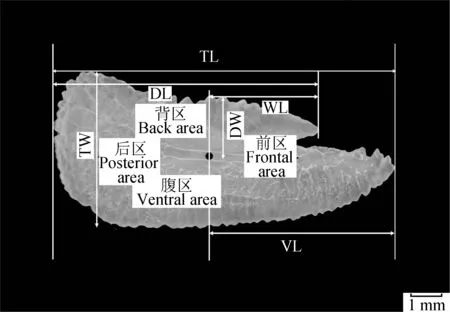
图3 蓝点马鲛耳石形态参数示意[61]Fig.4 Schematic otolith morphological parameters of Japanese Spanish mackerel S. niphoniusTL.耳石长; TW.耳石宽; DL.背长; DW.背宽; VL.腹喙长; WL.基叶长.TL.total length; TW.total width; DL.dorsal length; DW.dorsal width; VL.ventral length; WL.wing length.
4.2 体长及体质量生长
蓝点马鲛早期生活史可分为鱼卵、仔稚鱼和成鱼3个阶段[60]。其中仔稚鱼个体划分为卵黄囊期、前弯曲期、弯曲期、后弯曲期和稚鱼期5个发育阶段[61]。宋超等[62]对象山港鱼卵、仔稚鱼的时空分布特征及其与环境因子关系的研究表明,鱼卵、仔稚鱼与表层水温均呈显著性相关,表层水温为15~19 ℃时鱼卵丰度较高,仔稚鱼在表层水温为18~19 ℃时出现较多,而温度过高或过低时鱼卵和仔鱼丰度均较低。中国沿岸蓝点马鲛目前有记录的最大年龄达11龄,最大叉长为2.4 m,最大体质量为122.5 kg[63]。20世纪60年代初期, 蓝点马鲛繁殖群体平均叉长560~590 mm;20世纪80年代末期[22],平均叉长为466~598 mm;20世纪90年代[64],平均叉长为574 mm;20世纪初期(2006—2008年)[65],叉长为582 mm。近几年研究发现,繁殖群体(2016—2017年)平均叉长仅为554 mm[31]。总体趋势显示,近年来蓝点马鲛小型化现象有所加剧。目前蓝点马鲛生长过程一般使用von Bertalanffy方程描述。Shoji等[66]研究认为,蓝点马鲛的生长模型为:m=2.115×10-4L3.587。
5 繁殖习性
蓝点马鲛鱼卵的丰度与水深呈显著性相关,蓝点马鲛产卵群体倾向于在较深水域产卵[67]。楼丹等[34]研究发现,蓝点马鲛的产卵高峰时间为4月中旬至5月初。其主要产卵场有兄弟岛—厦门近海产卵场、牛山产卵场、闽东产卵场、浙江近海产卵场、吕泗产卵场、海州湾产卵场及日本的濑户内海产卵场(表2)。
蓝点马鲛个体绝对繁殖力在不同年龄阶段存在较大差异,1龄和2龄个体的绝对繁殖力较低,其个体绝对繁殖力为(9.2~25.9)万粒/尾,随着个体增长,3龄、4龄个体的绝对繁殖力增大至(19.8~81.8)万粒/尾,5龄、6龄个体的的个体绝对繁殖力高,为(16.6~127.5)万粒/尾,但7龄后个体的绝对繁殖力有所下降。20世纪90年代黄渤海蓝点马鲛样本的平均绝对繁殖力为97.8万粒/尾[68],2006—2008年其平均绝对繁殖力为68.0万粒/尾[38],而近年来其平均绝对繁殖力仅为50.5万粒/尾,可以发现蓝点马鲛绝对繁殖力逐年减小。此外,各年龄组(1龄除外)的绝对繁殖力、相对繁殖力也远低于历史水平,且随着年龄的增长其下降幅度增大。而繁殖力的变化与营养条件的改变有一定关系,营养条件恶化时,鱼类的绝对繁殖力就会减少[31]。
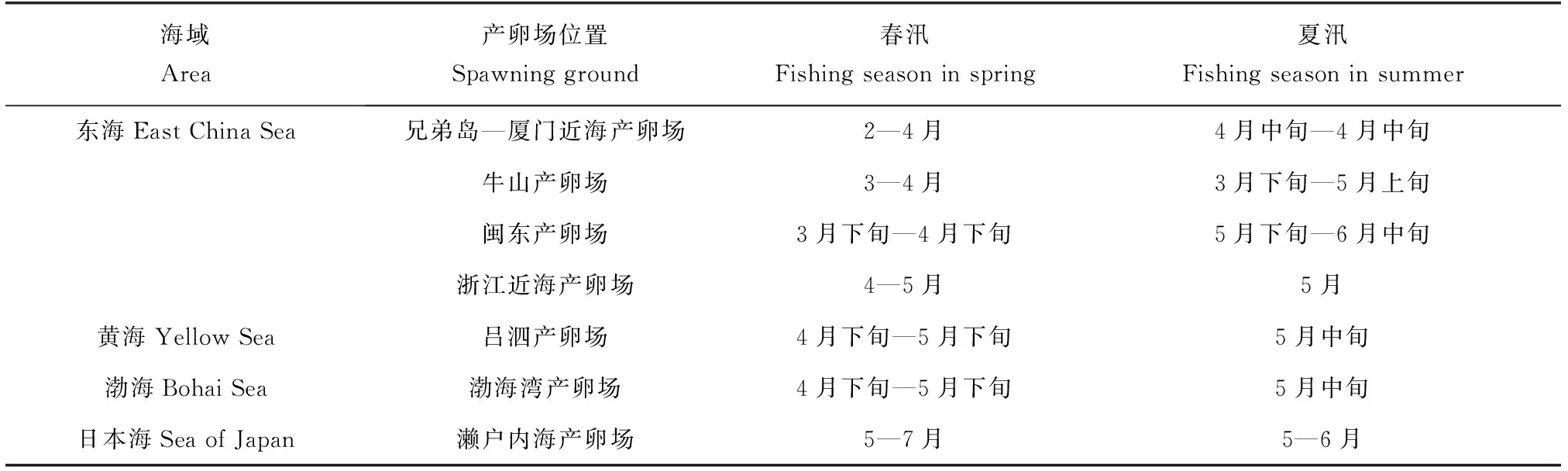
表2 蓝点马鲛主要产卵场分布[34,67]
6 摄食生态
鱼类早期个体的摄食能力和饵料丰度是影响个体生长和存活的重要因素[69-70],也是其能否成功进入到补充群体的关键所在[71]。饵料生物的缺乏,将使机体处于饥饿状态,导致生长速度降低,进而影响个体存活率。
在海洋生态系统中,蓝点马鲛处于食物链的中间位置,具有重要地位,既是海鸟以及海洋哺乳动物的重要捕食对象,又是主动的捕食者,主要捕食鳀鱼(Engraulisjaponicus)、玉筋鱼(Ammodytespersonatus)和细螯虾(Leptochelagracilis)等。林楠等[29]通过对象山港蓝点马鲛仔稚鱼肠道分析,发现蓝点马鲛仔稚鱼的饵料生物有11种,主要为鱼类仔鱼,鱼卵、磷虾、桡足类等也少量出现。也有学者研究了黄、渤海蓝点马鲛的摄食习性,研究表明其主要以鱼类为食,其次是虾类,主要饵料生物为玉筋鱼、细螯虾、日本鳀、中国毛虾(Aceteschinensis)[31]。相较于20世纪80年代,日本鳀作为蓝点马鲛的饵料生物占绝对优势地位[72-74];目前日本鳀虽仍为蓝点马鲛的主要饵料生物,但其优势地位已被玉筋鱼和细螯虾所替代[29],摄食结构发生变化,营养级下降。
7 分析与展望
7.1 种群结构
蓝点马鲛的种群结构复杂,目前仍无定论,但学者们普遍认为可将蓝点马鲛划分为黄海及渤海种群、东海种群。然而,种群内部可能会有亚种群的存在,每个区域的蓝点马鲛群体又有可能来自不同的产卵场、越冬场及索饵场,其种群关系如何依然不清楚,这些问题对我们合理开发、管理与保护该资源造成了一定的障碍。因此,黄海及渤海种群、东海种群的关联性是蓝点马鲛种群研究中有待解决的关键环节。未来对蓝点马鲛种群研究的试验个体样本采集,需注重时间和空间域上的广度(包含蓝点马鲛产卵场、索饵场及越冬场海域),以掌握蓝点马鲛群体分布、密度和季节变化。在此基础上,采用生物地球化学手段,“破译”耳石中保存的不同生活史阶段来自于生活环境的元素及稳定同位素“指纹”,以此确认其资源的时空关联性,从而更准确地划分蓝点马鲛地理种群。
7.2 年龄与生长
国内外学者对蓝点马鲛耳石的年龄生长已有较为完善的研究,目前蓝点马鲛耳石长与体长的关系、年龄结构等已有较为广泛的研究。但近年来蓝点马鲛小型化和低龄化现象愈发严重,未来还需从以下几个方面开展工作:(1)结合深度学习(卷积神经网络模型)等方法,对不同生长阶段蓝点马鲛群体进行识别,探究其生长阶段是否存在变化剧烈的“拐点”,并结合环境因子等,研究其“拐点”出现的原因。(2)完善蓝点马鲛日轮的鉴定,目前日轮的鉴定工作尚存诸多问题,如幼鱼耳石的处理方式、首轮形成的时间、产卵时间与孵化时间的逆推算。
7.3 摄食与繁殖
国内外学者对蓝点马鲛摄食生态与繁殖已有了初步的研究,相较于20世纪80年代,蓝点马鲛的主要饵料生物已从日本鳀替换为玉筋鱼和细螯虾,摄食结构发生变化,营养级下降,小型化、低龄化现象加剧,产卵期提前,繁殖力下降。近年来,蓝点马鲛摄食生态的研究主要为胃含物分析,常会过高估计食物中目标种的出现频率。随着稳定同位素技术的不断发展,未来可以将胃含物分析法与组织的稳定同位素法相结合,并利用脂肪酸分析以及特定化合物稳定同位素分析来研究蓝点马鲛长期的食性、食物的来源,确定蓝点马鲛在食物链中的营养层级。
7.4 资源量评估与管理
我国渤海、黄海、东海海域,与韩国和日本都存在专属经济区重叠的情况,因此对我国近海海域渔业资源的利用造成了很大的障碍[75-77]。能够理清在东、黄海蓝点马鲛的种群之间的关系和洄游路线变化,估算出相应的资源量,并且提出相对应的管理措施,可切实为我国争取到应得的渔业权益。此外,海洋灾害事件频发,导致近海生态结构不断变化,捕食者数量减少,鱼类种群数量发生变化等,蓝点马鲛资源特性发生相应改变,因此针对资源评估参数的估算是亟待解决的问题,在今后的研究中需进一步分析。
