尼古丁成瘾机制的研究进展
2021-07-19马晓丽
焦 瑒,贾 颖,朱 苹,张 倩,马晓丽*
(1.山东大学附属济南市中心医院 基础医学研究中心,山东 济南 250013;2.泰安市中心医院 检验科,山东 泰安 271000)
烟碱型乙酰胆碱受体(nicotinic acetylcholine receptors, nAChRs)属于配体门控离子受体家族,通过改变阳离子通道(Na+/Ca2+/K+)通量介导快速突触传递[1]。nAChRs是五聚体结构, 由5个跨膜单元组成,由9种α亚单位亚型(α2~α10)和3种β亚单位亚型(β2~β4)组成,以异聚体或同聚体的形式排列。尼古丁(nicotinic,NIC)是烟草的主要成分,其通过烟碱型乙酰胆碱受体(nAChRs)在脑中起作用。本文综述了nAChRs与NIC成瘾相关性的研究进展。
1 nAchRs亚型参与中脑边缘系统和尼古丁强化的调控
中脑边缘多巴胺(dopamine, DA)系统在行为激活或增强中起关键作用[2]。尼古丁暴露导致DA从腹侧被盖区(ventral tegmental area, VTA)释放到伏隔核(nucleus accumbent, NAC)。尼古丁是烟草的成瘾成分,通过nAChRs发挥作用[1]。中枢DA系统,特别是VTA神经元释放的DA在成瘾中起核心作用。
DA神经元以不同的组合表达含有α4、α5、α6、β2和β3亚基的异构化nAChRs,主要为α4/β2/α5和α4/α6/β2/β3[3]。α4β2-nAChR在尼古丁对认知、注意力和情感状态中起重要作用,是介导尼古丁成瘾的受体[4],其是目前已批准的戒烟药物的主要靶点之一。含有α6-nAChR在与愉悦、奖赏和情绪控制相关的中脑神经元中高度集中性分布,其主要分布于中脑DA神经元、蓝斑去甲肾上腺素(norepinephrine, NE)能神经元和视网膜神经节细胞,提示在调节中脑边缘功能中具有重要作用。DA神经元突触后去极化对尼古丁奖赏和强化行为(如尼古丁位置偏好和自我给药)至关重要[5]。α7-nAChR介导尼古丁诱导的VTA DA神经元的爆发性放电[6],α4β2和α7-nAChR不同程度参与尼古丁依赖和烟草成瘾过程。
DA神经元突触后去极化对尼古丁奖赏和强化行为(如尼古丁位置偏好和自我给药)至关重要。α-芋螺毒素TxIB作为α6/α3β2β3-nAChR (α6β2-nAChR)特异性拮抗剂,可降低尼古丁诱导的小鼠不同脑区如NAC、海马(hippocampus, HIP)和前额叶皮质(prefrontal cortex, PFC)中DA、GABA和NE的浓度,且VTA中DA和NE含量降低,γ-氨基丁酸(γ-aminobut-yric acid, GABA)变化不大。α4和α6-nAChR在NAC调节DA传递过程中,发挥重要作用。
2 与尼古丁厌恶和戒断有关的烟碱受体和通路
缰核(habenula, Hb)是一种上皮细胞结构,分化为内侧缰核(medial habenula, MHb)和外侧缰核(lateral habenula, LHb)两个核复合体[7]。MHb和LHb在解剖学、化学和功能上均属不同的亚核,每个亚核有不同的传入及传出神经元。LHb接收来自中脑和后脑的传入信息,并广泛投射到中脑和后脑,其中密集投射到喙侧被盖核(rostromedial tegmental nucleus, RMTg)。缰核复合体输出信号到中脑核团,在病理行为调节中起关键作用。损毁LHb可显著逆转尼古丁诱导的焦虑。MHb的下2/3由腹内侧、腹中央和腹外侧亚核组成[8],其主要是表达神经肽物质的神经元,接受来自前连合床核的输入,并同侧投射到脚间核(interpeduncular nucleus, IPN)的最外侧部分。IPN是一个非同源的大脑结构,它被细分为3个不成对的脚间亚核[顶端(apical sub-nucleus, IPA)、嘴侧(rostral sub-nucleus, IPR)和中央(central sub-nucleus, IPC)]和4个成对的亚核[背外侧(dorsolateral sub-nucleus, IPDL)、背内侧(dorsomedial sub-nucleus, IPDM)、外侧(lateral sub-nucleus, IPL)和中间(intermediate sub-nucleus, IPI)]。MHb主要由产生神经递质乙酰胆碱(acetylcholine, ACh)或P物质的神经元组成[8]。约90%-100%的MHb神经元表达α3、α4、α5、β2和β4-nAChR。内侧缰核-脚间核(medial habenula-interpeduncular nucleus, MHb-IPN)通路调节尼古丁戒断所产生的负性情感状态,该通路中的nAChRs拮抗作用可引起慢性尼古丁暴露小鼠的戒断样行为,提示尼古丁依赖信号的敏感性增高[9]。因此,nAChRs在MHb-IPN通路尼古丁厌恶和戒断中起重要作用(图1)。
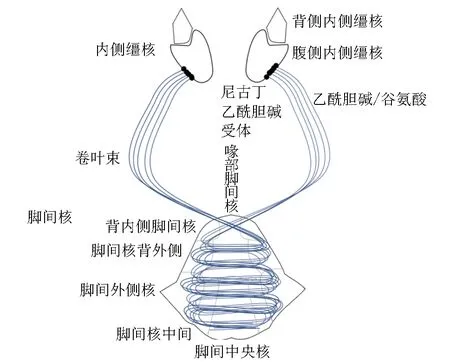
图1 nAchRs内侧缰核-脚间核轴尼古丁厌恶和戒断通路
2.1 α4β2-nAChR在尼古丁厌恶与戒断中的作用
α6-nAChR激活可以去极化LHb细胞,增加放电及LHb中谷氨酸的释放,α4β2-nAChR激活向LHb的GABA能输入,与α6-nAChR相比具有更快的脱敏作用[10]。在尼古丁依赖动物中,MHb神经元中α4β2-nAChR的上调是尼古丁戒断引起的焦虑增加的关键介质[11]。接受NIC注射的小鼠的脑放射自显像显示,在NIC戒断过程中,伏隔核壳、内侧缰核、丘脑核、背外侧膝状核、反曲束、腹侧被盖区、脚间核和上丘的α4β2-nAChR标记明显增加。尼古丁慢性治疗可增加脑内α4β2-nAChR位点,减少α6β2-nAChR位点,对α3β4-和α7-nAChR群体影响甚微。
2.2 其他nAChRs在尼古丁厌恶与戒断中的作用
α3和β4-nAChR在腹侧内侧缰核中广泛存在,α6、β2、β3和α4亚基选择性地存在于腹侧内侧缰核的部分区域[12]。MHb-IPN通路高度表达由CHRNA5-A3-B4基因簇编码的α5,α3和β4-nAChR[8],在该通路中,β4-nAChR在高尼古丁剂量下限制VTA-ICSA。α5-nAChR在MHb-IPN途径中表达最密集,同时也在许多其他成瘾相关的脑区中表达;如在VTA中,高表达α5-nAChR是滥用药物通过DA能神经元产生奖励和成瘾特性的基础;其在确定DA激活和尼古丁强化所需的最低尼古丁剂量中起关键作用。尼古丁通过刺激α5-nAChR激活MHb-IPN通路,nACh诱导的MHb-IPN通路激活发出负性动机信号,从而限制尼古丁的进一步摄入。
慢性尼古丁暴露期间,MHb-IPN通路失调是尼古丁戒断行为的基础。长期接触尼古丁后停止吸烟会引发戒断综合征,这是戒烟成功率低的重要原因[11]。戒断症状主要为出汗、心动过缓或胃肠不适和涉及焦虑和抑郁情绪的情感症状和认知缺陷。MHb胆碱能或谷氨酸能神经元中α6α4-nAChR信号上调是尼古丁戒断期间焦虑加剧的基础。在尼古丁依赖小鼠的MHb中输注α6-nACh选择性拮抗剂可以缓解戒断期间的焦虑样行为。上述研究结果支持α5和β4,可能还有α4β2-nAChR和α6-nAChR在调节尼古丁成瘾和尼古丁戒断综合征表达的MHb-IPN通路中的重要作用。因此,含有这些亚基的nAChRs可能是开发新的戒烟疗法的重要靶点。
除了MHb-IPN途径外,α5、α3和β4-nAChR亚基在后脑孤束核(nucleus tractus solitarii, NTS)中也高表达[13]。尼古丁激活了NTS中的胰高血糖素样肽-1 (glucagon-like peptide-1, GLP-1)神经元。GLP-1神经元表达α5-nAChR,其可以作为尼古丁的“饱腹感传感器”,刺激缰核系统,在尼古丁产生厌恶效应之前促进尼古丁的回避。尼古丁诱导的小鼠中脑边缘DA系统的激活受GLP-1受体调节;尼古丁等成瘾过程直接通过GLP-1受体介导。GLP-1受体是G蛋白偶联受体,能增强环磷酸腺苷(cyclic adenosine monophosphate, cAMP)的产生。在胰腺β细胞中,GLP-1刺激cAMP增加可导致核移位和β-连环蛋白的磷酸化,并使β-连环蛋白与转录因子7样2 (transcription factor 7-like 2, TCF7L2)二聚化[14]。TCF7L2是GLP-1信号级联的核心成分,在MHb神经元中高度富集[14]。TCF7L2在GLP-1受体介导的传递下游起作用,调节缰核nAChRs的功能,控制尼古丁的摄入。
3 尼古丁对抑郁、食欲和注意力的影响
尼古丁强化和避免尼古丁戒断的厌恶效应是尼古丁成瘾的基础,其他因素也可能导致人类的吸烟行为。严重抑郁障碍(major depressive disorder, MDD)和尼古丁依赖(nicotine dependence, ND)经常并存[15]。nAChRs激动剂可以改善动物模型中的抑郁行为,改善抑郁个体的情绪;吸烟可以调节焦虑症患者的症状严重程度,改变与恐惧和焦虑相关的行为,尼古丁对焦虑症的发展、维持和复发起重要作用。
3.1 nAChRs对焦虑和抑郁样行为的影响
中枢神经系统中的α7-nAChR在调节与抑郁症相关的小胶质细胞功能和神经炎性反应中起重要作用[16]。激活α7-nAChR有效地阻止了脂多糖(lipopolysaccharide, LPS)诱导的焦虑、认知障碍和抑郁样行为,并调节了HIP和PFC中相关的神经炎性标志物。尼古丁对抑郁样行为的影响也很复杂。在抑郁症患者中,β2-nAChR的有效性与终生抑郁发作次数、创伤评分和焦虑评分显著相关。在所有民族中,尼古丁依赖与抑郁情绪呈正相关。尼古丁依赖与抑郁障碍共病存在共同的途径或共同的诱发因素,如可能导致尼古丁成瘾和抑郁的遗传或环境因素。刺激尼古丁胆碱能受体可释放DA、谷氨酸和γ-氨基丁酸,从而影响神经性适应的发展并增高促肾上腺皮质激素释放因子的水平。这些变化可能在戒烟中起关键作用。
在与人类焦虑和抑郁相关的应激诱导行为中存在不同的nAChRs亚型[17]。通过乙酰胆碱酯酶(acetylcholinesterase, AChE)的局部敲除增加HIP中的乙酰胆碱信号,导致增加对焦虑环境的回避,在不可避免的压力下,不动性增加,社交失败的门槛降低。HIP中α7-nAChR的局部敲除极大地减弱了由AChE阻断引起的焦虑和社会相互作用[18],表明ACh-血清素相互作用可导致通过该结构介导的ACh信号的焦虑效应。HIP内ACh信号的上调通过nAChRs引起焦虑源性和社会交往障碍,nAChRs亚型调节基底外侧杏仁核(basolateral amygdala, BLA)的基线活动,降低其活性可诱导应激反应。Ach与去甲肾上腺素相互作用是调节BLA活性和相关的应激诱导行为关键之一[19]。ACh信号增加与注意力和学习能力的提高最为相关。ACh的最佳水平有助于奖赏性刺激和厌恶性刺激的编码。
3.2 nAChRs对注意力相关行为的影响
研究表明,精神分裂症患者比普通人群吸烟更多[20]。精神分裂症患者尼古丁自我用药改善认知症状,可能是通过促进尼古丁诱导的HIP α7-nAChR激活。尼古丁可以暂时改善精神分裂症患者的持续注意力,但尼古丁对精神分裂症相关的注意力受损没有明显的逆转作用。α7和β2-nAChR的失调是精神分裂症认知和情感症状的原因,尼古丁的使用可能是纠正这些症状的一种策略。除了尼古丁对成年期注意功能的影响外,nAChRs在发育过程中对注意力重要回路的成熟起作用[21]。DA系统的成熟促使青春期脑区域之间的功能连,有助于执行功能和认知控制的发展。突触富集的膜结合蛋白Lypd6是一种nAChRs功能调节因子,可以与人脑中的多种nAChRs亚型结合,是脑中胆碱能信号的多功能抑制剂,并在发育早期受到尼古丁暴露的调节。
3.3 nAChRs对食物摄入的影响
尼古丁对行为的表面敏感化效应部分反映了对其短暂抑制运动效应的耐受性;低剂量提高了食物和食物相关刺激的奖赏价值;高剂量增强了肌肉活动[22]。在大鼠中,自我给予尼古丁可以抑制体重,且不依赖于食物的摄入量。自我服用尼古丁即使在非常低的剂量下也会抑制体质量,减少尼古丁剂量会导致体质量增加。尼古丁通过改变脑中含有食欲肽和厌食肽的神经元的活性来影响能量平衡和食物消耗[23]。尼古丁通过nAChRs起作用,其中α3β4、α7和α4β2-nAChR与尼古丁对体重的调节有关。
厌食性阿片黑素促皮质激素原(pro-opiomelanocortin, POMC)神经元是哺乳动物脑中最著名的食欲抑制细胞[23]。尼古丁或胞嘧啶的给药激活了下丘脑弓状核(arcuate nucleus, ARC)中的POMC神经元,尼古丁诱导的α3β4-nAChR激活,使其与POMC神经元和黑皮质素系统相互作用来调节体重。尼古丁增加 ARC中促食欲激素相关肽(agouti related peptide, AgRP)和POMC神经元的放电率[24]。综上所述,除了通过POMC神经元信号传导对饱腹感产生影响外,中脑边缘系统中的乙酰胆碱也可能影响寻找可口食物的动机且通过不同的nAChRs亚型调节他们的享乐价值。
4 问题与展望
综述可见,nAChRs在脑部尼古丁成瘾中发挥关键作用(图2)。高亲和力α4β2-nAChR在尼古丁相关成瘾、情绪、注意力和食物消耗等行为活动中起关键作用。α4α6β2β3-nAChR及α5、α3和β4-nAChR在尼古丁强化、烟草依赖中起重要作用。此外,α3β4、α7和α4β2-nAChR在食欲调节中也起着重要作用,特别是尼古丁对食欲的抑制作用。进一步研究nAChRs亚型对尼古丁成瘾相关特性的作用及作用机制,将为烟草依赖新疗法的开发提供重要的新靶点。此外,这种新型疗法还可用于治疗情绪和注意力障碍以及控制体质量。
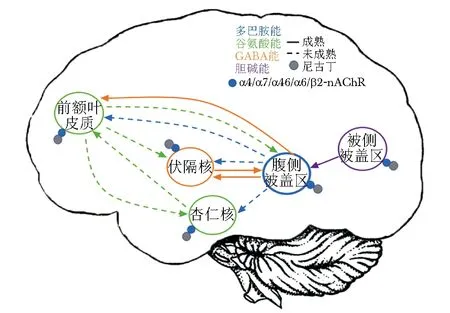
图2 nAChRs介导尼古丁成瘾神经通路
