围栏封育对珠芽蓼繁殖策略的影响
2021-07-19潘黎梅孟丽华
潘黎梅,孟丽华
(云南师范大学生命科学学院,教育部生物能源持续开发利用工程研究中心,云南省能源与环境生物技术重点实验室,云南 昆明 650092)
植物资源分配研究主要包括两个方面的内容:一是资源在地下部分和地上部分的分配,二是资源在生长和繁殖之间的分配.植物在生长过程中投入到繁殖器官的资源比例即繁殖分配,主要体现在植物对有限资源如何权衡生长与繁殖的分配及其在不同物种/生境中的变化规律.对繁殖分配的研究是植物生活史研究的核心内容之一,繁殖分配的变化是植物对多变环境适应的形式之一[1-3].对高山植物繁殖分配的研究多集中在揭示海拔梯度的变化对植物生长和繁殖权衡关系的影响.随海拔升高,植物生活的环境通常更加严酷,而在高海拔地区生长的植物通常会降低对营养繁殖和繁殖的资源分配而导致植株个体变小,但繁殖分配的比例却呈增加的趋势[4],而多数研究的结果也证明了这一点.例如,禾叶风毛菊[5](Saussureagraminea)和羌塘雪兔子[6](Saussureawellbyi)个体大小随海拔升高而降低,但繁殖分配则随着海拔的升高而升高.这些研究结果表明,在胁迫生境中植物将更多的资源投资到繁殖,表明繁殖对植物具有更为重要的意义.
植物除了投资繁殖的资源分配受环境影响外,植物的繁殖模式(有性繁殖和无性繁殖)通常也对环境变化表现出较强的规律性.珠芽蓼是一种兼具有性繁殖(花)和无性繁殖(珠芽)的植物,范邓妹等[7]调查了珠芽蓼两个海拔相似但生境不同的居群,结果发现在恶劣生境中珠芽蓼增加了对有性繁殖(花)投资的资源比例,相似的结果在不同海拔的珠芽蓼种群也得到了验证,随海拔升高珠芽蓼投资到花的资源比例呈增加的趋势[8].这些研究结果一方面表明恶劣环境中的有性繁殖比无性繁殖对植物具有更为重要的意义,另一方面表明生境变化会改变植物的资源分配模式.近年来,由于气候变化以及人类活动等外界因素的影响,生境的破坏、退化等生态环境问题日益严峻,加强对脆弱生态系统的保护已经引起了广泛关注并付诸实施.在草地生态系统保护中,围栏封育作为一种有效的措施得到了广泛地应用.围栏封育后,围栏外的草地会比围栏内的草地受到更多的干扰,而且围栏内的生物多样性(包括植物多样性、微生物多样性和土壤动物多样性)会呈现增加的趋势[9-11],但对植物多样性的影响与封育的时间密切相关.显然,围栏封育后导致围栏内外的生境产生了较大的差异,而这种生境差异如何影响植物的繁殖模式仍然缺乏研究.因此,本研究通过比较围栏内外珠芽蓼的繁殖分配策略,以期理解植物的繁殖策略对围栏封育的反应.
珠芽蓼(PolygonumviviparumL.)是蓼科(Polygonaceae)蓼属(Polygonum)多年生草本植物,在北半球陆地广泛分布.珠芽蓼具有无性繁殖(珠芽)和有性繁殖(花)两种繁殖方式,已有的研究表明珠芽蓼的有性繁殖和无性繁殖比例在不同生境中存在显著差异,而海拔、温度、营养[7,12]和微环境[13]都能影响有性繁殖和无性繁殖的资源分配比例,表明珠芽蓼是研究生境异质性对繁殖策略影响的理想材料.适度的围栏封育能有效地改善区域环境[14],使围栏内外生境发生改变.因此,研究围栏内和围栏外植物的繁殖行为差异能较为准确地反应围栏封育对植物繁殖策略的影响.本研究比较了高寒草甸生态系统中围栏内外珠芽蓼的植株特征和投资到有性繁殖和无性繁殖的资源比例变化,揭示围栏封育通过改善区域的生态环境进而对植物的资源分配产生影响.本研究重点回答以下问题:1)围栏封育对珠芽蓼的植物特征有什么影响? 2)围栏封育对珠芽蓼有性繁殖和无性繁殖的资源配置模式有什么影响?
1 材料和方法
1.1 研究地概况
本研究的野外采样于2019年7月在中国科学院海北高寒草甸生态系统定位研究站(以下简称海北站,N 37°37′,E 101°19′,海拔3 200 m)附近的高寒草甸进行.海北站位于青藏高原的东北部,祁连山北支冷龙岭东段南麓的大通河河谷.海北站的气候为典型的高原大陆性季风气候,区域年平均气温为-1.7 ℃,年均降水量约为580 mm[15];海北站周围的主要植被类型为青藏高原典型的地带性植被,以金露梅(Potentillafruticosa)为建群种的高寒灌丛草甸和以嵩草属(Kobresia)植物为建群种的高寒嵩草草甸[15].海北站已经建立了长期围栏封育永久样地用于开展高寒草甸生态系统的检测等试验,永久样地的围栏外为当地牧民的草地牧场,周边在夏季经常作为牧场使用.围栏内的采样地点位于海北站永久样地内,而围栏外的采样地点在永久样地的旁边.两个样地均为矮嵩草(K.humilis)草甸,常见植物为垂穗披碱草(Elymusnutans)、异针茅(Stipaaliena)、紫羊茅(Festucaovina)、珠芽蓼、圆穗蓼(Polygonummacrophyllum)、麻花艽(Gentianastraminea)、湿生扁蕾(Gentianopsispaludosa)等.
1.2 试验材料
珠芽蓼为多年生草本植物,广泛分布于海拔1 200~5 100 m的亚高山和高山草甸湿润地区[17],在我国西北、华北、东北、西南、中部地区均有分布.珠芽蓼的花序呈穗状,上面着生珠芽和花,其中珠芽可在母株上萌发叶芽,珠芽成熟后自然脱落入土,生根并生长为新植株[18];花序上的花可通过传粉产生种子,但结籽率通常很低;珠芽蓼的花数量和珠芽数量可以作为投资到有性繁殖和无性繁殖的资源状态.可依据花序上着生的繁殖器官可以将植株分为三类.一类是花序上同时着生有性繁殖的花和无性繁殖的珠芽,第二类花序上全部着生花,第三类是全部着生珠芽,但后两种类型所占比例很小[17].
1.3 试验方法
在围栏内和围栏外分别随机选择40个植株,用卷尺测量每个植株从基部到花序顶部的高度,然后采集每个植株的地上部分并单独装在信封内.室内统计每个植株上的花数量、珠芽数量、花和珠芽总数,并用花数量/(花数量+珠芽数量)计算每个植株上的花所占的比例.
1.4 统计分析
围栏内和围栏外的植株高度、花数量和珠芽数量、珠芽和花总数量以及花所占比例的差异用独立样本t检验(independent-Sample T Tests)比较.利用回归分析(regression analysis)统计植株高度与花数量、珠芽数量、花和珠芽总数量、花所占比例的关系,以及花数量和珠芽数量的关系.所有数据用SPSS 25.0统计软件包进行比较分析.
2 结果与分析
围栏内珠芽蓼的植株高度(t=4.384,P<0.05)和珠芽数量(t=4.038,P<0.05)均显著高于围栏外的植株(图1),而花数量(t=-3.644,P<0.05)却显著低于围栏外的珠芽蓼植株(图1).珠芽和花总数量在围栏内外不存在显著性差异(t=-0.127,P>0.05),然而围栏外的花所占比例(t=-4.633,P<0.05)却显著高于围栏内的植株(图1).
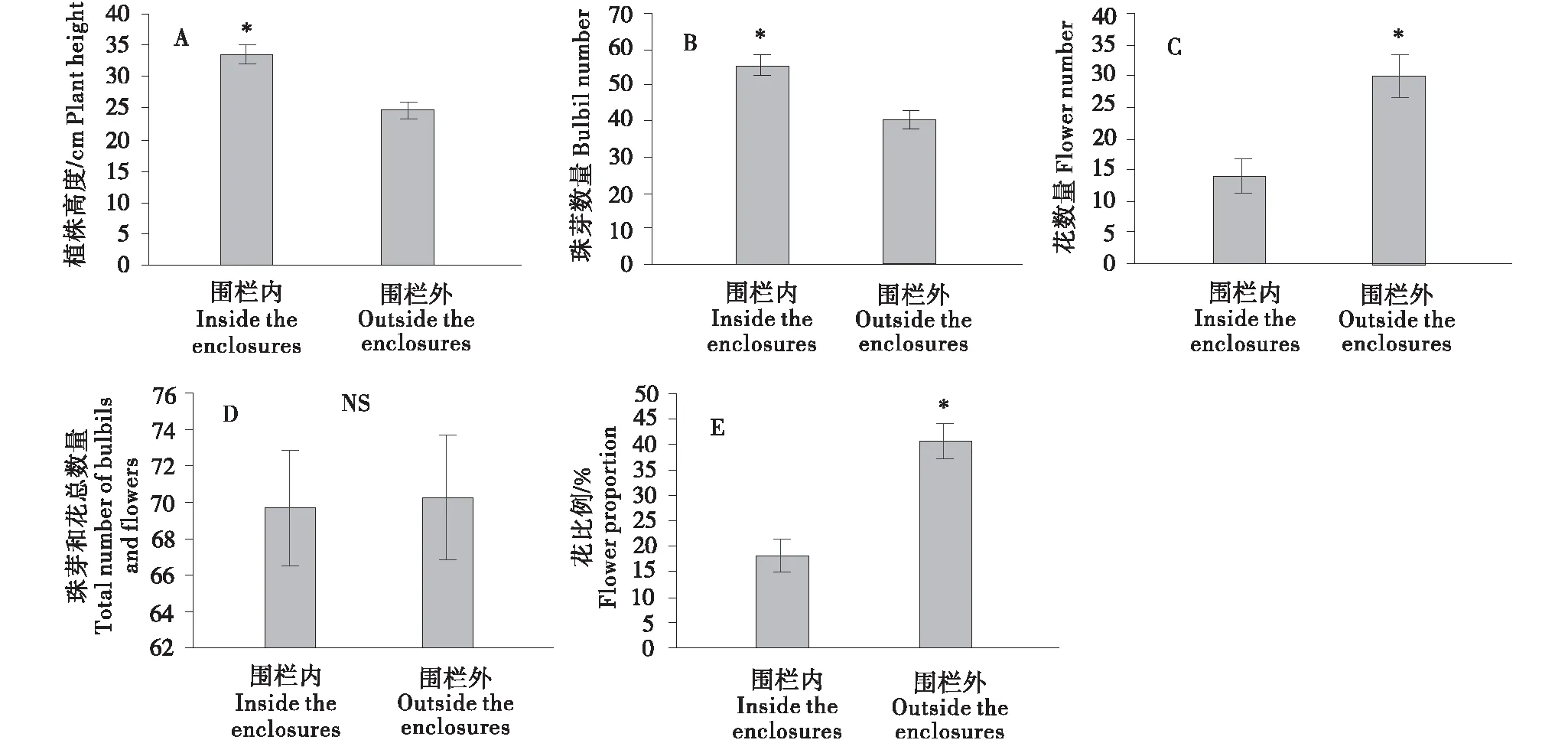
* 表示在 0.05 水平差异显著,NS 表示在 0.05 水平差异不显著.
珠芽蓼的植株高度和珠芽数量之间在围栏内(r=0.534,P<0.05)和围栏外(r=0.424,P<0.05)都存在显著的正相关关系(图2-A),而植株高度和花数量在围栏内(r=-0.121,P>0.05)和围栏外(r=-0.074,P>0.05)存在不显著的负相关关系(图2-B).受围栏内外珠芽数量和花数量不同变化趋势的影响,围栏内珠芽蓼的植株高度与珠芽和花总数量(r=0.386,P<0.05)存在显著的正相关关系,但在围栏外(r=0.225,P>0.05)存在不显著的正相关关系(图2-C).珠芽蓼的植株高度与花所占比例在围栏内(r=-0.256,P>0.05)和围栏外(r=-0.249,P>0.05)存在负相关关系,但均不显著(图2-D).
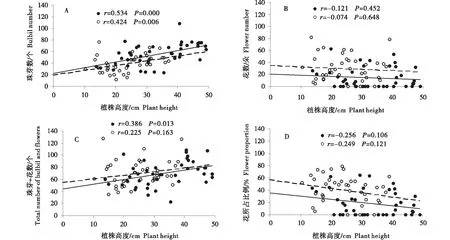
图2 围栏内(黑色点)和围栏外(白色点)珠芽蓼植株高度与花数量(A)、珠芽数量(B)、花和珠芽总数量(C)、花所占比例(D)的关系
对围栏内外每个植株的花数量和珠芽数量进行回归分析,结果表明围栏内(图3-A,r=-0.358,P<0.05)和围栏外(图3-B,R=-0.366,P<0.05)植株上花和珠芽的数量均呈现显著的负相关关系.

图3 珠芽数和花数量在围栏内(A)和围栏外(B)的关系
3 讨论
围栏封育可以有效改善草原的区域生态环境[18],并能在很大程度上改变植物群落结构和组成.通常来说,围栏内的植被因缺乏动物啃食,禾草类植物的高度会显著增加[19],这会增加群落的郁闭度而引起其他植物的高度增加以获取更多的光资源.相比之下,围栏外的禾草类植物由于经常被牛羊啃食而导致植株高度降低,而同一群落的其他不可食植物高度则无需增加植株高度以竞争光资源;此外,围栏封育会改善土壤质地和提高土壤肥力[20-21],受这些因素的综合影响,围栏外植物物种的高度通常会低于围栏内的植物物种.本研究对珠芽蓼的研究结果进一步证明了这一观点,围栏内的珠芽蓼个体的植株高度显著高于围栏外,表明围栏封育会增加围栏内植物的生物量和平均高度,说明适度的围栏封育对提高牧草产量具有非常重要的作用.
围栏内植株的高度增加表明植物所获得的资源也发生了变化,因此植物对繁殖的投入也会发生相应的改变.植物个体越大,对繁殖资源的投入越高;但植物对繁殖器官的投入增加后,需要额外增加对营养器官的投入以支持植物的繁殖过程及其支撑结构,因此植物对营养器官生长的投入和对繁殖器官的投入通常存在异速生长关系,导致较大的植物对繁殖资源投入的比例降低[4-5,22-23].对珠芽蓼的研究结果表明,围栏内植株高度显著高于围栏外的植株,围栏内植株无性繁殖体(珠芽)的数量显著高于围栏外的植株(图1-B),而植株高度与珠芽数量显著的正相关关系同样证明了较高的珠芽蓼会产生较多的珠芽数量(图2-B).这些结果证明珠芽蓼植株的资源状态会直接影响珠芽蓼的无性繁殖体数量.有性繁殖器官(花)的结果与营养繁殖器官(珠芽)表现出相反的趋势,围栏内珠芽蓼的花数量和花所占繁殖器官的比例均显著低于围栏外的珠芽蓼植株.已有的结果表明,在环境更为胁迫的环境中珠芽蓼会增加对花资源的投资[7],而胁迫生境中的植物同样会增加对有性繁殖投资的比例[7,15,24-25].正如以上指出,围栏外的种群容易受到动物践踏、土壤肥力较差等因素的影响,本研究的结果进一步证明珠芽蓼在胁迫的环境中会增加对有性繁殖器官资源比例的投资,表明有性繁殖对植物具有更为重要的意义.
植物的无性繁殖模式会促进植物开拓新生境并迅速建立种群,但通过无性繁殖方式建立种群的遗传一致性高,对潜在环境变化的应对能力差[26].与之相对应,有性繁殖会促进植物的基因交流而增加种群的遗传多样性[27],但有性繁殖(特别是异花传粉的交配系统)通常需要传粉媒介(特别是生物媒介)的协助,因此对环境的要求较高.在资源有限的前提下,兼具有性繁殖和无性繁殖的植物对两种繁殖方式的投资通常呈现出负相关关系,即“此消彼长”的权衡关系[7].本研究对珠芽蓼围栏内外种群的研究结果进一步证明了有性繁殖和无性繁殖的权衡关系.
致谢:感谢中国科学院海北高寒草甸生态系统定位研究站全体工作人员给予本研究工作的帮助!
