低温冷藏对卷丹百合珠芽呼吸强度与萌发生根的影响
2023-10-27王军利千小绵孙越赟冯树林
王军利, 千小绵, 周 佳, 孙越赟, 李 乐, 高 潮, 冯树林
(1.咸阳职业技术学院, 陕西 咸阳 712046; 2.陕西省农业宣传信息中心, 陕西 西安 710003; 3.陕西省现代农业园区服务中心, 陕西 西安 710003; 4.陕西秦草生态环境科技有限公司, 陕西 西咸 712046; 5.陕西省三原县农业科学技术中心, 陕西 三原 713800)
0 引言
【研究意义】卷丹百合(LiliumlancifoliumThunb)为百合科(Liliaceae)百合属(Lilium)常见球根花卉,在我国野生分布范围广泛,其生境主要为荒坡草地、灌丛、林下和岩石缝隙[1]。卷丹百合植物花期7—8月,花朵硕大,花色艳丽,生长健壮,喜温暖,耐贫瘠,喜干燥气候,不耐水涝,具有较强的抗寒、抗旱能力,能对地面形成长期的覆盖,具有极高的园林价值;其地下茎富含淀粉等营养物质,食用价值较高;同时,卷丹百合还具有较高的药用价值[2-5]。卷丹百合为三倍体,高度自交不育,其繁殖方法主要有珠芽繁殖、鳞片扦插繁殖、鳞茎自然分割繁殖及种球繁殖等4种。因长期球茎或鳞片繁殖会造成种性退化,同时,珠芽繁殖简单方便,且能保持种的优良性状,因此,卷丹百合在生产上以珠芽繁殖为主[6-8]。卷丹百合的珠芽是其地上茎叶腋部位产生的气生小鳞茎[9]。所以,开展卷丹百合珠芽繁殖的相关研究对促进相关产业发展具有重要现实意义。【前人研究进展】目前,对于珠芽发育和生理变化方面的研究较多[6, 10-13]。樊金瓶等[6]研究了卷丹百合珠芽发育形态特征及生理变化。张慧等[11]研究赤霉素(GA3)及其抑制剂(PAC)对卷丹和珠芽生长的影响发现,喷施GA3使花期提前15 d左右,并促进卷丹株高、花苞长和宽及节间长分别较对照(蒸馏水)增长69.73%、29.15%、37.16%及58.81%;喷施PAC则使花期推迟18 d,并使卷丹上述指标相应减小77.06%、30.91%、8.93%及69.86%,增加植株茎粗;GA3抑制珠芽形成从而使单株数量减少,但可增加单个珠芽重量;PAC则显著增加珠芽厚度并且珠芽分化效率最高(56.27%)。刘玉平等[12]研究发现,喷施10 μmol/L独脚金内酯(GR-24)及其抑制剂(Tis-108)对卷丹珠芽生长发育有明显影响,GR-24对珠芽发生有抑制作用,Tis-108则有促进作用;Tis-108处理卷丹珠芽直径和珠芽数均高于CK和GR-24。张慧等[11]研究发现,GA3虽抑制珠芽形成但可使母株上的珠芽不经休眠直接萌发。何长芳等[14]研究发现,随春化时间增加,同一地区卷丹的发芽率和生长势呈下降趋势,卷丹在45 d左右时发芽率与生长势最好;而相同春化时间下,不同地区卷丹的发芽率和生长势存在较大差异。呼吸作用是指植物器官或组织在酶的催化作用下,将其本身贮藏的营养物质进行一系列氧化分解,释放CO2、H2O及能量的过程。呼吸作用的强弱一般用呼吸强度表示,反映器官或组织代谢的快慢及生长的强弱,可以用作衡量植物器官或组织休眠、萌发或萌动的一项指标[10]。一般情况下,休眠植物或器官的呼吸强度较弱,而完成春化后,其呼吸强度急剧升高,使得萌发所需要的中间物质及能量得到充分供应。【研究切入点】低温有打破休眠,完成春化的作用[14-15]。目前,有关珠芽萌发和育苗方面的研究相对较少,特别在打破珠芽休眠、促使其度过春化及早萌发的机理及实践方面的研究鲜见报道。【拟解决的关键问题】以秦岭北麓汤峪及岱峪中采集的卷丹百合珠芽为研究对象,采用单因素试验,通过比较卷丹百合珠芽在不同冷藏温度(4 ℃、8 ℃及12 ℃)条件下珠芽生长及萌根率、呼吸速率及萌发生根等指标的变化规律,探索温度处理对其呼吸强度和形态发育等方面的影响,以期筛选其破除休眠、完成春化作用的适宜冷藏温度与处理时间,为卷丹百合产业化发展提供参考。
1 材料与方法
1.1 材料
1.1.1 百合珠芽 卷丹百合珠芽于2022年7月上旬采自秦岭北麓岱峪和汤峪山间居民的房前屋后。此时为卷丹的盛花期,采集每株茎上叶腋处所生的全部大、小珠芽,采获总量为4.6 kg。
1.1.2 仪器设备 DW-25W147型可调温冰箱,武汉医捷迅安商贸有限公司生产;LI-6262型CO2/H2O分析仪,美国LI-COR公司生产。
1.2 方法
1.2.1 不同贮藏条件对珠芽生长的影响 试验设4个处理,即室温〔(25±2) ℃,T0(CK)〕、4 ℃(T1)、8 ℃(T2)和12 ℃(T3)。将珠芽平均分成10份,分别装入容积为1 L的大烧杯;再将其中9个大烧杯平分成3组,分别放入设定温度的冰箱中冷藏,烧杯口用单层湿纱布覆盖;另外1个烧杯放实验室室温保存,烧杯口用双层湿纱布覆盖。每3~4 d将烧杯从冰箱中取出,揭开纱布,轻轻震摇、翻搅其中的珠芽,使其获得均匀的氧气及低温环境。然后,再将烧杯覆盖单层湿纱布,放回原处理环境;室温处理相同。在整个试验过程中,观察和记录珠芽的颜色、硬度、光亮度、健康程度及根的萌发与否、根的颜色、健康程度等表观特征。
1.2.2 不同冷藏条件对珠芽呼吸强度和根生长的影响 分别在处理后0 d、10 d、20 d、30 d、40 d、50 d和60 d时,从各处理的3个烧杯中随机取样,每个烧杯取样1次分别测定相关指标,取3次平均值。
1) 呼吸速率。参照刘亭等[16]的方法,称取各处理的珠芽50 g,将其放在对应处理温度下的密闭容器中,接通预热好的仪器。待仪器显示的数字变化平稳时,记录CO2浓度,并开始计时,5 min时再次记录CO2浓度,并计算珠芽的呼吸速率。其中,0 d数据的测取,是在未分装的混合珠芽中随机称取50 g,用上述方法测得其呼吸强度。
2) 根生长。于各处理的3个烧杯中分别随机取出100粒珠芽,统计发根珠芽数;用游标卡尺测量萌发根的长度,统计萌发根的总数,然后计算各处理不同时段珠芽的萌根率(A,%)、平均根长(L,mm)、平均根数(N,根/株)及不同冷藏条件下各时段珠芽的萌根率增幅(G,%);同时,观察并记录样本的外观状态,即珠芽及根的萎蔫程度、根的色泽及健康状况等。
A=生根珠芽数/总珠芽数×100%
L=∑根长/总根数
N=总根数/生根珠芽数
G=A1-A0
式中,A0和A1分别为上一测试时间点的平均萌根率和下一测试时间点的平均萌根率。
1.3 数据统计与分析
采用Excel 2019进行数据统计和制图,采用SPSS 19.0对数据进行对比分析。
2 结果与分析
2.1 不同冷藏条件下珠芽的生长
2.1.1 珠芽及根的表观特征 由表1看出,0 d时(新采)各处理的珠芽新鲜、光亮、较硬且均无根;此后,随处理时间延长,不同处理珠芽及根的表观特征变化均存在不同程度的差异。在3种冷藏温度(T1~T3)下,珠芽均慢慢发根,而在室温〔T0(CK)〕中放置的珠芽,直到最后(60 d时)样品中仍无根萌发。
处理0~10 d:各处理的珠芽均新鲜、光亮且硬,其中,T0和T1均一直未萌发根,仅T2和T3在处理10 d时萌发有少量白色且健壮的单根。处理20 d:T0的珠芽光鲜度开始降低且稍变软,但未萌发根;T1~T3的珠芽均新鲜、光亮且硬,其中,T1有少量白色健壮的单根萌发,T2和T3有白色且健壮的单根及少量双根萌发。处理30 d:T0珠芽光鲜度开始降低且稍变软,并未萌发根;T1~T3的珠芽均新鲜、光亮且硬,其中,T1有少量单根及双根萌发,T2有单根及双根萌发,T3有较多单根及双根萌发。处理40 d:T0的珠芽色泽开始降低且稍变软,但未萌发根;T1和T2的珠芽均新鲜、光亮、硬且萌发有较多白色健壮的单根及双根;T3的珠芽光亮度降低、变软,其有单根及双根萌发,白色,但根尖微黄。处理50 d:T0的珠芽色泽变暗、略皱缩且稍变软,仍未萌发根;T1的珠芽新鲜、光亮、硬且萌发较多白色健壮的单根及双根;T2的珠芽光泽度降低且变软,萌发白色健壮的单根及双根;T3的珠芽萎蔫、起皱、变软,其有少量单根及双根萌发,但根尖微黄、回勾。处理60 d:T0的珠芽色泽变暗、略皱缩且稍变软,仍未萌发根;T1的珠芽色泽变暗、稍皱缩且变软,有白色健壮单根及双根萌发;T2的珠芽略萎蔫、皱缩且变软,有单根和少量双根萌发,但有部分根根尖发黄、回勾。T3的珠芽萎蔫、皱缩且变软,有少量单根及双根萌发,但根弱且细,多呈黄褐色,根尖回勾且变干。综合看,T1(4 ℃冷藏条件下)珠芽在40~50 d时发根较快、较多,40~60 d时珠芽活力较强;T2(8 ℃冷藏条件下)珠芽在40~50 d时发根较多,珠芽活力充沛,其后活力逐渐变弱;T3(12 ℃冷藏条件下)珠芽发根较早,但变软、变皱也较早。说明,高温促使珠芽休眠,而低温冷藏有利于珠芽保鲜,虽发根较晚但发根较多,其根保持白色及健康较持久。所以,在珠芽萌根的相关数据统计分析中,未统计或列入室温下各处理天数的相关数据。
2.1.2 珠芽萌根率 由表2看出,盛花期时所采珠芽均无根;随着培养时间延长,各处理珠芽的发根率均呈上升趋势,增幅呈先升后降趋势;但不同贮藏条件下珠芽的萌根率和增幅存在差异。
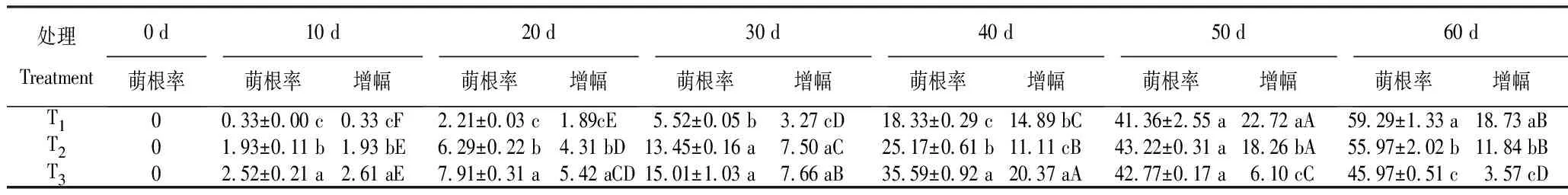
表2 不同温度下珠芽的萌根率随贮藏时间的变化
1) 萌根率。各处理前40 d珠芽的萌根率差异均显著,均依次为T3>T2>T1。其中,10~20 d时各处理珠芽的萌根率均<8%;30 d时T2及T3珠芽的萌根率均>13%,分别为13.45%和15.01%,显著高于T1;40 d时各处理珠芽的萌根率为18.33%~35.59%;50 d时各处理的萌根率为41.36%~43.22%,依次为T2>T3>T1,差异不显著;60 d时各处理的萌根率为45.97%~59.29%,依次T1>T2>T3,差异显著,与40 d时的结果相反。表明,处理40 d后,T1的萌根率增长较快,而T3减慢。
2) 萌根率增幅。萌根率增幅能够显示在某一阶段珠芽萌动的速度与活力。T3珠芽萌动相对较早,其萌根率增幅在30~40 d时达峰值,随后便显著下降;T2萌根率增幅在40~50 d时达峰值,其后显著下降。T1珠芽萌根最晚,萌根率增幅在40~50 d时达峰值,其后显著下降。处理后前40 d,每一时段的萌根率增幅均以T3最为显著;但40 d后,每一时段的萌根率增幅则以T1最为显著。表明,处理40 d后,T1珠芽快速萌动,而T3及T2珠芽的萌动速度降低,活力减退。
2.2 不同冷藏条件下珠芽的呼吸强度
由图1可见,常温状态下卷丹珠芽的呼吸速率未见明显峰值,且呈缓慢下降趋势。可能与温度过高不利其萌动有关;且随着时间延续及珠芽水分散失,其休眠逐渐加深。低温冷藏条件下,珠芽的呼吸速率较之于常温状态均有显著峰值。因为低温能够打破珠芽的休眠,增强呼吸作用,使珠芽萌动生根[14-15]。T3珠芽的呼吸速率随贮藏时间延长而呈较快升高趋势,至30 d时达峰值,为208 mg/(kg·h),随后呼吸强度显著下降;T2和T1珠芽的呼吸速率峰分别出现在40 d和50 d,分别为263 mg/(kg·h)和259 mg/(kg·h)其后,随着贮藏时间延续,其呼吸速率均显著下降。60 d试验时段内3个冷藏温度下呼吸速率均出现峰值,且峰值之后呼吸速率下降。表明,3种冷藏温度下,60 d内卷丹珠芽均完成了春花作用,并开始萌发。呼吸速率峰值出现后下降时,春化作用才完成。冷藏温度分别为4 ℃、8 ℃和12 ℃时,其春化完成的时间分别大致为55 d、50 d和40 d。
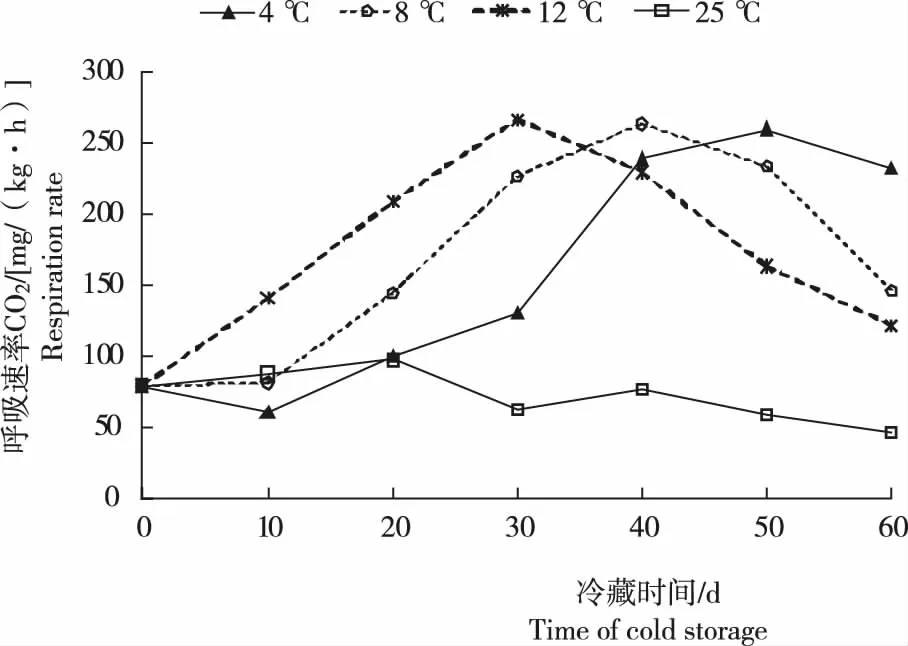
图1 不同贮藏温度及时间处理珠芽的呼吸速率
2.3 不同冷藏条件下卷丹珠芽的根长、根数及最大根长
从表3看出,不同冷藏条件下卷丹珠芽的平均根长、株平均根数及最大根长存在差异。

表3 不同冷藏温度及时间处理珠芽的根长、根数及最大根长
2.3.1 根长 不同冷藏处理各时段珠芽的平均根长差异显著。同一时段,10~30 d时T3和T2的平均根长显著长于T1;40 d时各处理间差异显著;50 d时各处理间无显著差异;60 d时T1显著长于T3和T2。表明,前期冷藏温度较高的珠芽先发育,但后劲不足,而冷藏温度较低时,发育推迟,但后劲强劲。平均根长在3个温度处理中均随时间延长而增长,同一处理各时段间差异显著或不显著,说明在60 d试验时段内,先期萌生时根长较短,随着时间的推移,其持续生长;同时,中后期萌生的根生长较快。
2.3.2 根数 同一时段冷藏处理前期不同处理间发根珠芽的株平均根数均无显著差异;中、后期各处理间差异显著或不显著,珠芽株平均根数的最大值分别出现在T3(30 d、40 d)及T1(50 d、60 d)。同一处理,T1的珠芽株平均根数50 d显著高于其余时段,60 d与50 d差异显著,且均显著高于其余处理;T2的50 d显著高于其余时段,60 d与40 d差异不显著,但均显著高于其余处理;T3的株平均根数40 d显著高于其余时段,30 d与50 d差异不显著,但均显著高于其余处理。原因在于双根现象(1个珠芽生2个根)在不同处理中的不均等概率造成。前期发根较少,且萌生的新根生以单根为多;中、后期时,各处理由于冷藏温度不同,对珠芽萌发的春化作用大小不同,新根生长获得的物质和能量在不同时期存在显著差异,从而造成有的处理中珠芽生出较多双根,而同时期其他处理中双根较少。
2.3.3 最大根长 冷藏试验前期,珠芽萌生根的最大根长出现在T3(10 d、20 d、40 d)及T2(30 d),后期则出现在T2(60 d)及T1(50 d)。同一时段除40 d时的最大根长T3与T2差异不显著外,其余时段各处理间差异显著;同一处理除T1的20 d与30 d差异不显著外,其余处理不同时段间差异均显著。与上述结果相互对应,即前期T3及T2的珠芽较早启动发育,而后期T2及T1的珠芽萌发后劲较大。
3 讨论
在夏、秋季节,部分卷丹百合的珠芽于形成后在茎秆的叶腋间持续发育,并最终萌生少量根。大部分珠芽在还未萌出根时便从植株上掉落,掉落的珠芽在地表受到高温、失水及病虫害等威胁。进入冬季,珠芽经过较长的低温春化,在早春温度回升时萌发出土,其内在春化萌动过程在冬季的低温期完成[11, 14-15]。低温冷藏有利于打破休眠、完成春化,使植物整体或其器官提前萌发[15],从而改变因春化而受限休眠植物的播种期。前人的试验较少涉及打破卷丹百合珠芽休眠使其完成春化的具体温度[13-14]。试验结果表明,12 ℃即可打破卷丹百合珠芽休眠;打破卷丹百合珠芽休眠使其完成春化的时间为50 d左右,且该温度对珠芽内部生理代谢活动的抑制作用不强。冷藏温度为8 ℃和4 ℃时也能满足卷丹百合珠芽的春化要求,但随着冷藏温度的降低,珠芽内部前期的生理活动受到的抑制较大;后期,随着珠芽春化作用的完成,其内部的生理代谢过程很快加强,珠芽大量萌根,且能较持久保持健康和活力。观察发现,珠芽在贮藏期间其形态变化、萌根率及萌根率增幅均与其各自的呼吸速率存在明显的对应关系:萌根率增幅最大值总是出现在呼吸速率峰值出现及以后的一段时间。可见,呼吸作用为珠芽根的萌发提供了物质和能量,是萌发的前提,此结果与植物生理学中春化作用的过程及机理[15]相互印证。说明,植物或植物器官打破休眠完成春化通常是温度、时间、水分供应及通气状况等多因子综合作用的结果[15]。研究仅测定了温度和时间对卷丹百合珠芽春化作用的影响,但呼吸速率的变化显示则是多因子作用后的综合表现结果。何长芳等[14]研究认为,卷丹百合珠芽最佳春化时间为45 d,其后,随着春化时间的延长,珠芽的发芽率和生长势不断下降。本研究结果表明,较多双根出现的时段(平均根数>1.25根)与其处理中珠芽的呼吸速率峰值高度关联,其总是在呼吸速率峰值的同期及稍后时段出现。春化完成后,随着春化时间的延长,其发芽率及长势不断下降的原因是珠芽呼吸速率降低、根发黄变褐、根尖回勾变干及珠芽生命力变弱等造成。说明,呼吸作用强为珠芽提供萌发的物质和能量多,使双根出现的几率增大;呼吸作用较弱双根出现的概率随之减少[15]。与何长芳等[14]的研究结果基本一致并相互印证。
试验证明,夏秋季常温条件下卷丹百合的珠芽处于休眠状态,其以较弱的呼吸维持生命活力,保持体内营养物质;冬季低温条件下,珠芽完成春化,继而在春季萌发出土。4~12 ℃的低温冷藏可打破卷丹百合珠芽休眠,促使其完成春化并萌发,但在不同冷藏温度条件下,呼吸速率峰值的出现时间不同,从而使植物保持旺盛萌发活力的时效及时间点不同;呼吸速率最大及稍后一段时间是植物打破休眠后的最适播种期,此结论与植物生理学原理相符[15]。但依据植物生理学春化作用的原理,春化积累刚完成之后,随即给予较高温度有解除春化的作用。所以,在应用春化作用打破休眠使植物提早萌发时,应注意避免在春化作用刚完成时使植物处于高温环境中,继而使春化效果被削弱甚至解除[15]。
4 结论
室温〔(25±2) ℃〕贮藏及低温(4 ℃、8 ℃和12 ℃)冷藏处理,卷丹百合珠芽的呼吸速率、萌发生根及珠芽外观表现等随贮藏时间变化而变化,其中,温度越高,珠芽萎蔫越快,活力下降越早;室温条件下贮藏的珠芽一直不萌发生根,其珠芽的呼吸速率呈缓慢下降趋势;4 ℃、8 ℃和12 ℃冷藏条件下,珠芽的呼吸速率均呈先升后降趋势,表明经过一段时间的低温处理后,卷丹百合的珠芽均能完成春化作用开始萌发。综合考虑各处理春化完成后珠芽的外观健康状态、活力、生根率及呼吸速率的保持等情况认为,打破休眠使其完成春化并开始萌发的最佳条件为冷藏温度4 ℃贮藏时间55 d。
