不同水分条件下小麦粒重QTL定位及其元分析
2021-07-19倪胜利何瑞刘媛张沛沛李兴茂杨德龙
倪胜利,何瑞,刘媛,张沛沛,李兴茂,杨德龙
(1.甘肃省农业科学院旱地农业研究所,甘肃 兰州 730070;2.甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
小麦(TriticumaestivumL.)是世界上主要的粮食作物之一,其产量占世界粮食总产量的30%以上.决定小麦产量的主要因素有单位面积穗数、穗粒数和粒重,各产量因素之间相互作用,共同决定小麦的产量.在穗数和穗粒数一定的条件下,粒重对小麦产量提高具有重要的作用[1-3].因此,研究小麦粒重遗传特性,对提高小麦粒重遗传改良效率具有重要的意义.
小麦粒重属于微效多基因控制的数量性状,易受环境影响[4].据前人研究,小麦粒重主要受加性效应控制[5-6].王瑞霞等[5]以和尚麦×豫8679为亲本的RIL群体为试验材料,定位了35个与粒重有关的QTL,分布在小麦1A、1B、2A、2D、3A、3B、4A、4D、5A、5B、6D和7D染色体上,可解释表型变异4.36%~16.80%;周淼平等[6]在多个环境下对小麦产量性状进行QTL定位时,检测到与粒重相关的QTL位点共5个,分别位于2A、2B、3B、4D和7A染色体上,单个QTL可解释9.6%~25.7%的表型变异;胡亮亮等[7]在多个环境不同水分条件下定位了19个与粒重相关的QTL位点,分布在除1A、3B、4D和6A以外的其他17条染色体上,并且发现了在多个环境下均能够稳定表达的QTL位点以及4个QTL热点区域Xmag2064-Xbarc181(1B)、Xwmc522-Xgwn122(2A)、Xwmc446-Xgwm610(4A)和Xwmc603-Xbarc195(7A).以上研究表明,控制小麦粒重的QTL位点在不同群体以及不同环境中表达数目和特点各不相同,这对小麦粒重相关QTL的准确定位造成了很大的困难.
元分析(meta-analysis)通过对不同群体和遗传背景中的QTL位点信息进行整合验证,确定一致性QTL(MQTL),进一步细化了QTL位置,减少了实验间的误差[8-11].目前,元分析已经广泛应用在作物QTL位点的整合研究中.Chardon等[8]通过对玉米花期相关性状的QTL位点进行元分析,发掘出62个MQTL,将玉米花期相关QTL的定位精度提高了两倍;吴琼等[9]通过对大豆生育期相关的QTL位点进行元分析,发现了7个与开花期相关的MQTL和2个与成熟期相关的MQTL;叶亚琼等[10]通过对311个不同群体中与小麦株高和粒重相关的QTL位点进行元分析,得到28个与株高相关的MQTL和21个与粒重相关的MQTL;Hanocq等[11]使用元分析方法得到2个控制早熟性基因和4个QTL热点区域.
本研究以陇鉴19和Q9086为亲本杂交创建的重组自交系群体(RIL)进行粒重性状分析,同时将定位到的QTL同收集到的QTL进行元分析,发掘“真实”QTL位点,为深入剖析小麦粒重的遗传基础和QTL精细定位奠定理论基础.
1 材料与方法
1.1 试验材料
以陇鉴19和Q9086为亲本杂交创建的RIL群体120个株系为供试材料.陇鉴19是甘肃省农业科学院旱地农业研究所选育的新品种,为甘肃省第一个大面积应用的抗旱北移冬小麦新品种,抗旱性强,耐瘠薄,对水肥反应不敏感.Q9086是西北农林科技大学农学院小麦研究所提供的品系,对水肥反应敏感,尤其灌浆期对水分反应敏感[12].
1.2 田间试验与性状测定
试验于2014年10月~2015年7月,2016年10月~2017年7月和2017年10月~2018年7月在甘肃省榆中县小麦试验点进行,地理坐标N 35°51′,E 104°07′,平均海拔1 900 m,平均气温6.6 ℃,年均降水量450 mm,年均蒸发量1 450 mm,无霜期140 d.3个环境依次记为E1、E2和E3.在每个环境中,整个生育期仅播前施基肥1次,基肥量均为纯钾 60 kg/hm2,纯氮180 kg/hm2,纯磷150 kg/hm2.田间试验采用随机区组设计,设干旱胁迫(drought stress,DS)和正常灌溉(well-watered,WW)两个水分处理,每个处理3次重复,行长1.0 m,行距0.2 m,每行点播60粒,每株系种植6行.具体水分管理措施为:播种前统一灌溉底墒水(900 m3/hm2),干旱胁迫处理仅在小麦拔节期灌水750 m3/hm2,之后完全依靠自然降水;灌溉处理则分别在小麦拔节期、抽穗期和开花期补充灌水,每次灌水量为750 m3/hm2.其中3个年度环境下小麦全生育期降水量依次分别为130、142和137 mm.籽粒完全成熟后,按株系分别收获和风干后,使用小麦籽粒自动考种分析仪(SC-G,杭州万深)对各株系千粒重(thousand grain weight,TGW)进行测定.
1.3 数据统计与QTL定位分析
对小麦RIL群体千粒重数据采用SPSS V21统计软件进行分析.广义遗传力(h2B)的计算按照Toker[13]提出的方法计算.
结合前期构建的小麦RIL群体分子遗传连锁图谱(共定位了524个SSR标记,形成21个连锁群,全长22 66.7 cM,两标记间的平均遗传距离是4.3 cM)[14-15]进行千粒重QTL定位分析.在QTL IciMapping V 4.0软件中采用复合区间作图法CIM(composite interval mapping)对3个环境不同处理下的RIL群体千粒重加性QTL(A-QTL)进行检测及遗传效应分析.
1.4 QTL数据收集与整理
通过Web of Science(http://isiknowledge.com)和中国知识资源公共数据库(http://www.cnki.net)查找并收集国内外发表的关于小麦千粒重相关QTL位点信息[8,16-30,33],根据BioMercator 4.2软件的标准对收集的QTL位点信息进行整理,包括QTL名称、染色体位置、置信区间、连锁系数、贡献率、侧翼标记和群体大小等[15].
1.5 QTL信息映射
以高密度遗传图谱wheat composite 2004(http//wheat.pw.usda.gov/ggpages/mapse shortlist.html.)为参考图谱,该图谱总长度为2 569 cM,共1 235个SSR标记,标记间平均距离为2.2 cM.在BioMercator 4.2软件中将收集的QTL位点信息数据和参考图谱进行整合,构建一致性图谱.通过高斯定理最大似然比计算一致性QTL可能存在的位置和置信区间,选取AIC(akaike-type criteria values,AIC)值最小化的模型为最佳模型,即“真实QTL”模型.
2 结果与分析
2.1 千粒重表型分析
对3个不同水分环境条件下,小麦RIL群体及其双亲TGW表型分析发现,正常灌溉条件下TGW的平均值(45.01~46.58 g)显著高于干旱胁迫条件下的平均值(37.1~42.96 g).但干旱胁迫条件下TGW的变异范围(21.20~50.28 g)大于正常灌溉(33.33~57.45 g),且干旱胁迫下TGW表型的变异系数(7.06%~13.60%)高于正常灌溉(5.95%~8.08%),说明干旱胁迫是影响小麦TGW的重要环境因素之一.从群体的分布情况来看,小麦RIL群体的TGW表现出超亲分离的现象,群体的分离近似于正态分布(图1).

E1~E3分别表示2014~2015、2016~2017和2017~2018年度试验点,每年度试验点均设干旱胁迫(DS,条纹柱表示)和正常灌溉(WW,空心柱表示)处理;R、L和Q分别表示RIL群体、陇鉴19和Q9086;Mean、Range、CV(%)、Skew.和Kurt.分别表示RIL群体千粒重表型平均值、变异范围,变异系数、偏度和峰度.
从表1中可以看出,TGW的表型变异受到水分环境(E)、基因型(G)以及基因型与环境之间互作(G×E)的显著影响(P≤0.01).其中,水分环境均方(9 758.30**~15 131.76**)显著大于基因型均方(27.52**~91.75**)和基因型与环境互作均方(7.26**~34.15**),说明水分环境是影响TGW的主要因素之一.通过对广义遗传力(h2B)的分析,在3个环境中小麦TGW的h2B为0.63~0.74,说明在水分环境之外,基因型也是控制小麦TGW表型变异的主要因素.
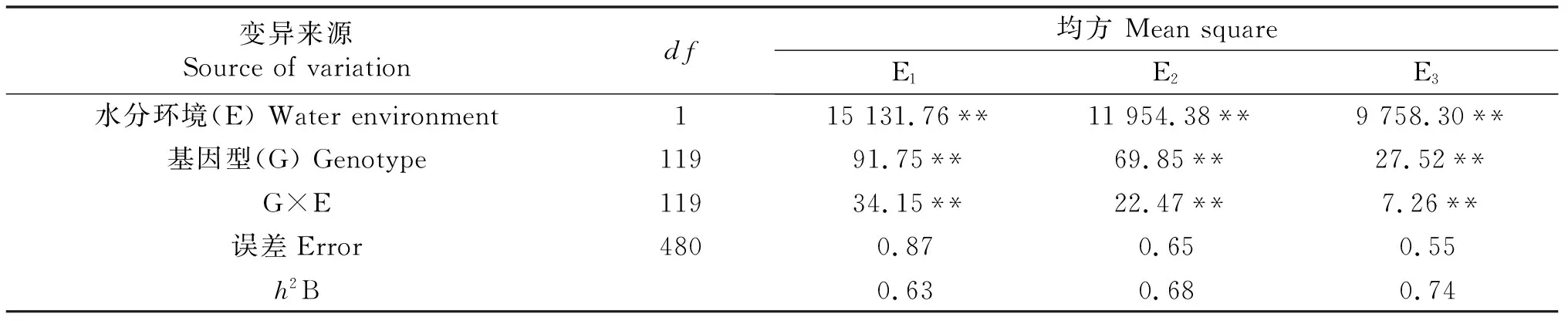
表1 小麦RIL群体千粒重方差分析
2.2 千粒质量QTL加性效应分析
从表2中可以看出,利用IciMapping V 4.0软件采用复合区间作图法,在不同处理条件下,共检测到14个控制TGW的加性QTL位点,分布在1B、3B、4B、4D、5B、6A和6B染色体上(表2).其中Qtgw.cas-1B.1、Qtgw.cas-3B.1、Qtgw.cas-4B.1、Qtgw.cas-4D.1、Qtgw.cas-4D.2、Qtgw.cas-5B.1和Qtgw.cas-6A 7个QTL位点具有降低TGW的效应,其加性效应(A)在0.62~1.32 g,贡献率(R2)在5.06%~12.15%;Qtgw.cas-3B.3和Qtgw.cas-6B两个QTL位点具有提高TGW的效应,A在1.01~1.04 g,R2在6.03%~6.46%.同时,在1B、3B、4B和5B上发现了5个在2个以上环境中均稳定表达的QTL位点,Qtgw.cas-1B.2(E1~E3)、Qtgw.cas-1B.3(E1,E3)、Qtgw.cas-3B.2(E2,E3)、Qtgw.cas-4B.2(E1~E3)和Qtgw.cas-5B.2(E1,E3),这些位点在不同的环境中均能够表达,但其对表型的作用方式在不同环境以及不同水分处理中有明显差异,说明这些位点对环境有着较强的适应性.
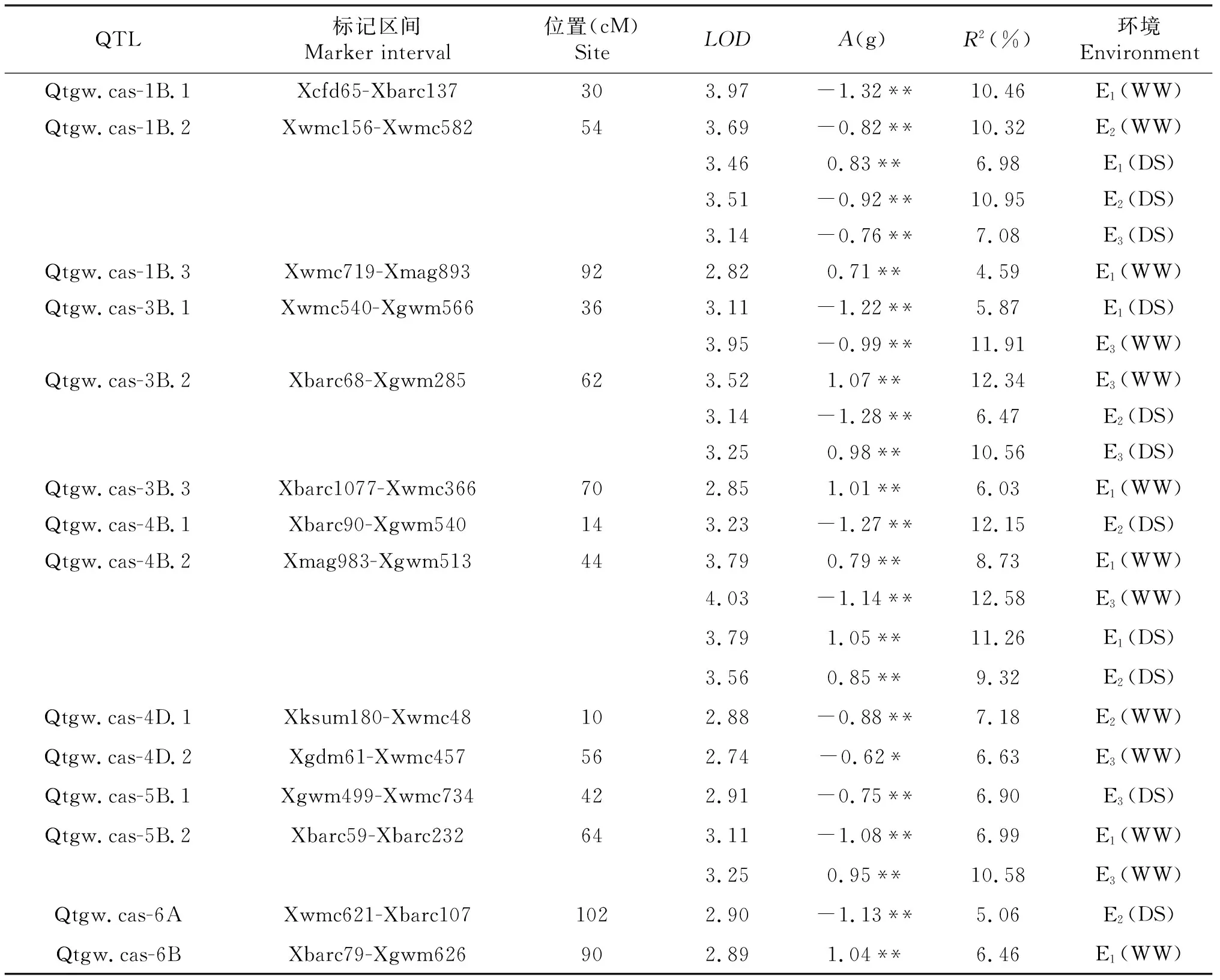
表2 不同水分环境条件下小麦RIL群体千粒重QTL加性效应及其对表型变异的贡献率
同时,小麦RIL群体在不同环境及不同处理下控制TGW的位点在不同染色体和同一染色体的不同区段上都呈不均匀分布(图2),在1B、3B和4B染色体上分布最多,达到5~7个,而在其余染色体上分布相对较少.这些QTL位点在对应的染色体上聚集分布,形成QTL热点区域,如1B染色体上Xwmc156-Xwmc582,3B染色体上Xbarc68-Xgwm285,4B染色体上Xmag983-Xgwm513.说明在这些染色体区段上可能存在控制TGW的重要基因.

三角形、圆圈和正方形分别表示2014~2015、2016~2017和2017~2018年甘肃省榆中县(N 35°51′,E 104°07′)试验点所检测到的控制小麦千粒重的QTL;空心图形表示正常灌溉条件下检测到的QTL,实心图形表示干旱胁迫条件下检测到的QTL.
2.3 QTL元分析
对来自14篇文献共13个不同作图群体及定位得到的155个粒重QTL(表3),通过元分析共得到38个一致性QTL(表4),主要分布在1B、2B、2D、3A、3B、3D、4A、4D、5B、6A、6B和7A染色体上,平均每条染色体上有3.1个MQTL.在这些MQTL中,有10个MQTL位点的置信区间小于4 cM,即MQTL2(0.95 cM)、MQTL3(0.46 cM)、MQTL15(2.04 cM)、MQTL17(3.08 cM)、MQTL27(2.2 cM)、MQTL30(1.7 cM)、MQTL31(0.95 cM)、MQTL33(1.45 cM)、MQTL37(3.51 cM)和MQTL38(1.94 cM).有3个MQTL的图距小于4 cM,即MQTL2(3.54 cM)、MQTL4(3.98 cM)、MQTL19(2.94 cM),同时发现MQTL30和MQTL31位于6A染色体相邻位置且图距均为7.00 cM,MQTL37和MQTL38位于7A染色体的相邻位置且图距均为6.67 cM.从图3中可以看出,MQTL在染色体上不均匀分布,在1B、2B和5B染色体上分布较为集中,可能在这些区段中包含有控制TGW的基因.

每条染色体左侧的横线代表QTL的LOD值;竖线代表QTL的置信区间.

表3 小麦千粒重QTL数据的整合
3 讨论
粒重是小麦产量构成的重要因素之一,是多基因控制的复杂数量性状,易受环境的影响[19].研究发现在干旱胁迫条件下,小麦的粒重随着干旱程度的增加而下降,同时其表型变异增大,遗传力下降,说明水分环境对小麦粒重有显著作用[29].本研究发现,与正常灌溉条件相比,干旱胁迫条件下小麦TGW平均下降了6.46 g,变异系数增加了3.59%.TGW表型变化趋势与前人研究结果相同,再次证明了干旱胁迫是小麦粒重的重要影响因素.前人研究表明,控制小麦TGW的QTL位点在小麦21条染色体上均有分布,但是在不同遗传背景不同环境中稳定表达的QTL位点的数目和效应不同[5-7,15-28].李美霞等[26]在波兰小麦1B、2A染色体上检测到了1个和2个与小麦粒重相关的QTL位点;周淼平等[6]在2A、2B、3B、4D和7A染色体上检测到5个控制TGW的QTL位点.本研究在1B、3B、4B、4D、5B、6A和6B染色体上共检测到14个与粒重相关的加位性QTL位点,其中Qtgw.cas-1B.2和Qtgw.cas-4B.2在多个环境中均能稳定表达.同时Qtgw.cas-1B.2在3个环境中干旱胁迫条件下稳定表达,并对TGW表型起负调控作用.在1B染色体上定位了很多与粒重相关的QTL位点,但是本研究中定位到的QTL位点与其他研究中的位点并未重合,属于新定位的QTL位点,还有待进一步研究.
同时,元分析通过将来自不同环境和作图群体中的QTL位点进行整合和优化,筛选出“一致性”QTL,为后续的精细定位和图位克隆奠定基础.近年来,元分析在玉米、大豆等作物改良中发挥了重要作用.江培顺等[30]利用元分析对玉米穗行数、行粒数和粒重相关的584个QTL位点进行整合,得到了22个穗行数MQTL、7个行粒数MQTL和 44个粒重MQTL,并进一步发掘得到了10个与玉米产量相关的基因以及12个与水稻产量相关基因具有同源性的候选基因;李长育等[31]对47个与大豆结瘤性状相关的QTL进行元分析得到2个MQTL,并通过基因注释得到8个与大豆结瘤相关的基因;Tyagi等[32]对80个与小麦籽粒性状相关的QTL位点进行元分析,在1B、2A、2D、3B、4A、5A、6A和6B染色体上确定了23个相关MQTL,并鉴定出3个与小麦粒重相关的重要MQTL.本研究收集整合了155个与粒重相关的QTL位点,通过元分析共得到38个一致性QTL,置信区间小于4 cM的MQTL有 10个,置信区间最小为0.46 cM,图距小于4 cM的MQTL有 3个,图距最小为2.94 cM.进一步说明元分析可以缩小QTL位点的置信区间,从而精确QTL的位置,提高作物分子育种和遗传改良的效率.
4 结论
通过QTL的定位分析,在不同的环境条件下共检测到14个控制千粒重的QTL位点,分布在1B、3B、4B、4D、5B、6A和6B染色体上,并在1B、3B、4B和5B染色体上发现了5个在2个以上环境中均稳定表达的QTL位点.通过对13个不同作图群体155个QTL位点进行元分析,共得到了38个MQTL,将置信区间缩小到0.46 cM,图距缩小到2.94 cM,进一步提高了QTL定位精度.同时,本研究发现在7A染色体Xfba204-Xfbb18b区段中存在两个紧密连锁的MQTL,说明在这个标记区间中可能存在控制TGW的“真实”QTL,尚需进一步研究证实.本研究通过对千粒重QTL位点进行定位以及在不同作图群体中进行元分析,确定了稳定表达的QTL及其QTL富集区域,缩小了置信区间,将为小麦粒重的QTL精细定位和分子标记辅助选择提供理论依据.
