红松枝叶关系的纬度差异性*
2021-07-16王明琦金光泽刘志理
王明琦 金光泽 刘志理
(东北林业大学生态研究中心 森林生态系统可持续经营教育部重点实验室 哈尔滨 150040)
植株选择性调整各组织相对生长速率可增强其适应不同环境的能力,提高自身竞争力(侯兆疆等, 2014; Özdenetal., 2018; Wangetal., 2018),同时这也是植物在整个生长过程中重要的生态学策略,可有效提升其资源获取和利用效率(Niklasetal., 2002; Lohieretal., 2014)。“枝叶大小关系”是植物生态学策略的主要维度之一(Niklasetal., 2002; Yanetal., 2013),反映了植物在水分传输和光拦截之间的适应性选择(Özdenetal., 2018),与植物碳水经济学密切相关(Westobyetal., 2002)。枝大小直接影响叶片分布(Canhametal., 1990; Posadaetal., 2012), 叶大小和数量直接决定植物冠层的紧密度,进而影响其暴露于光线下的程度(程徐冰等, 2011)以及碳捕获和光截取能力(Osadaetal., 2015; Fanetal., 2017; 李曼等, 2017)。顶部树枝是植物分支系统中最活跃的部分,也是随外部条件(如水和养分)变化最容易改变的部分,因此,研究当年生小枝的变化规律对深入了解植物生存策略、阐明植物对全球气候变化的响应机制至关重要(Westobyetal., 2002; Mengetal., 2015; Todaetal., 2018; Wangetal., 2019)。
“Corner’s rules”表明,枝横截面积较大的植物分枝较少,枝横截面积较大时其单叶面积也较大(White, 1983a; 1983b)。Smith等(2017)进一步研究“Corner’s rules”发现,单叶面积与总叶面积呈正相关。枝叶大小呈正相关,且为异速生长关系(Sunetal., 2006; Normandetal., 2008; Yangetal., 2009; 2014)。Sun等(2006)研究指出,即使生境压力并没有直接改变枝叶大小的异速生长指数,但随着海拔升高其异速常数(y轴截距)也会降低; Yan等(2013)研究表明,即使演替阶段并没有直接改变枝叶大小的异速生长指数,但其异速常数在不同演替阶段也存在显著差异。Brouat等(1998)研究发现,等速的生长关系亦存在于枝叶性状之间。目前,常用出叶强度逐步替代叶数量,即单位小枝体积或茎干质量的叶数量(Kleimanetal., 2007; Yangetal., 2008; Yanetal., 2013)。出叶强度和叶大小在不同坡向(史元春等, 2015)、生活型(Yangetal., 2008)以及草本植物(Whitmanetal., 2010)间存在等速生长关系,且为负相关; 异速生长关系与等速生长关系同时存在于被子植物体内(Scottetal., 2012); 不同海拔间,植株叶大小与出叶强度呈异速生长关系(李曼等, 2017)。 可见,物种和生境是影响枝叶大小和出叶强度关系的重要因素。
纬度不同带来的温度、降水差异,会造成植物所处生境以及光照有效性发生变化(Osadaetal., 2015),而针对不同生境,植物也会采取相应的适应机制(Sunetal., 2006; Mengetal., 2014)。如高纬度地区温度相对较低,植物会调整枝叶大小的形态以适应温度降低带来的枝木质部导管直径减小(Lietal., 2017)以及水和营养物质运输效率下降与叶温降低导致的代谢分解和光合羧化受损(Frenneetal., 2013)。Lusk等(2018),根据“叶能平衡假说”对温度变化带来的叶片大小等变异做出了解释,同时也强调了植株的温度敏感性。因此,探究同种植株枝叶性状在不同纬度的差异,对了解植株响应环境变化具有重要意义。
阔叶红松(Pinuskoraiensis)林是我国东北地区的地带性森林植被,是温带针阔混交林的典型代表(徐丽娜等, 2013)。本研究以沿纬度梯度从北到南在胜山、丰林、穆棱和长白山4个阔叶红松林分布区的主要建群种红松为研究对象,测定其当年生单叶面积、枝横截面积、出叶强度和总叶面积等枝叶性状,分析枝叶性状在不同纬度的差异,阐明枝叶性状关系在不同纬度之间的权衡策略。
1 研究区概况与研究方法
1.1 研究区概况
野外调查在我国东北阔叶红松林典型分布区进行,采样点从北到南(纬度由高到低)依次为黑龙江胜山国家级自然保护区、黑龙江丰林国家级自然保护区、黑龙江穆棱东北红豆杉(Taxuscuspidata)国家级自然保护区和吉林长白山国家级自然保护区,样地基本信息见表1。

表1 采样点基本信息Tab.1 Basic information of sample sites
1.2 数据采集
2017年7—8月,于4个不同纬度阔叶红松林分布区采样,每个采样点随机选取3株生长坡度相似的成年(胸径>45 cm)红松样树,共12株(表2)。为了减少个体树冠内部变异引起的枝叶性状对研究结果的影响,将冠层分为上、中和下3层,每层分为北和南2个方向并在每个方向选择1个相对完整的主枝。对于每个主枝,随机收集10个当年生小枝(第1个终端节点与分枝分离的部分到枝条末端),即每株样树60个样枝。保证样枝上叶片完整,密封(封口袋)带回实验室。样枝采集后6 h内,摘取样枝上所有针叶,测其平均针长(精度为0.01 cm)和体积(排水法,精度为0.01 cm3)。基于以下公式计算样枝总叶面积(A, cm2) (刘志理等, 2012):
(1)
式中:n为针叶数;v为样枝上支撑所有针叶的体积(cm3);l为平均针长(cm)。
单叶面积近似为平均叶面积,即总叶面积除以叶片数量; 样枝基径采用游标卡尺(精度为0.01 cm)测量。计算基径时,枝横截面近似为圆形; 计算出叶强度时,样枝为圆柱体,高近似为枝长,底面积近似为枝横截面积,得出枝体积后再计算单位枝体积支撑的叶片数量,即出叶强度(Yanetal., 2013)。

表2 采样点样本信息①Tab.2 Sample information of sample sites
1.3 统计分析
采用单因素方差分析-Tukey法检验4个采样点红松枝叶性状的差异,将其对数转化为正态分布, 使用嵌套方差分析法解析枝叶性状变异,即采用‘nlme’包的‘lme’函数对每个性状构建线性混合模型,其嵌套水平为采样点>个体>枝(Paradisetal., 2004)。
不同纬度的红松枝叶性状关系可表示为y=bxa,转化为lgy= lgb+algx,其中b表示枝叶性状关系的截距;x和y为特征参数;a为斜率,即异速生长参数或相对生长指数,当|a|显著偏离1时,二者为异速变化关系; 当|a|= 1时,二者为等速变化关系(Harveyetal., 1991)。采用MATR Version 2.0软件分析数据,所用方法为标准化主轴估计法(standardized major axis,SMA)。异速分析根据Warton等(2002)方法判定,即将回归斜率与|1|进行显著性检验。统计检验使用R-3.4.4进行。
2 结果与分析
2.1 红松枝叶性状的纬度差异
不同纬度红松枝叶性状差异显著(P<0.05) (表3)。枝横截面积的变异系数为33%~43%,穆棱采样点的枝横截面积均低于其他采样点。叶面积变化趋势为先增后减,不同纬度间的变异系数差异小,单叶面积和总叶面积的变异系数分别为20%~26%和66%~72%。随着纬度增加,出叶强度呈下降趋势,不同纬度间变异系数差异大,为69%~92%。
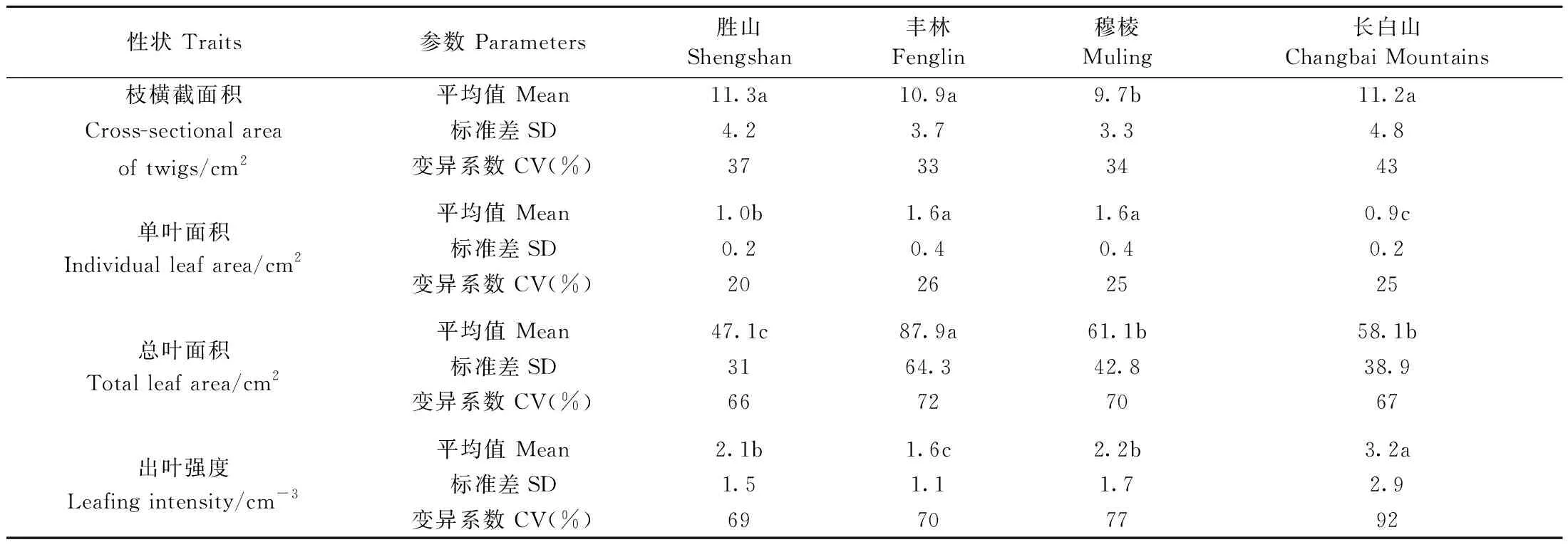
表3 红松枝叶性状的纬度差异①Tab.3 Difference of twig and leaf traits of P. koraiensis along latitude
枝叶性状变化的枝水平解释率普遍较高,其中对枝横截面积的解释率高达92% (图1),与表3分析结果一致,在穆棱采样点,各方向枝横截面积均较小(表4)。总体分析时其他采样点之间没有显著差异(表3),方向分析时差异显著(表4); 个体间枝叶性状差异小,个体水平对4种枝叶性状差异的解释率均低于18%, 枝横截面积、出叶强度和总叶面积的纬度解释率均小低于8%,但单叶面积变化的纬度解释率最高为58% (图1)。

图1 红松枝叶性状的嵌套方差分解Fig. 1 Nested variance decomposition of twig and leaf traits of P.koraiensisTCA: 枝横截面积; TLA: 总叶面积; ILA: 单叶面积; LIV: 出叶强度。 图中未显示即解释率小于0.1%。下同。TCA: Twig cross-sectional area; TLA: Total leaf area;ILA: Individual leaf area; LIV:Volume-based leafing intensity. If not shown in the figure, the explanatory is less than 0.1%. The same below.
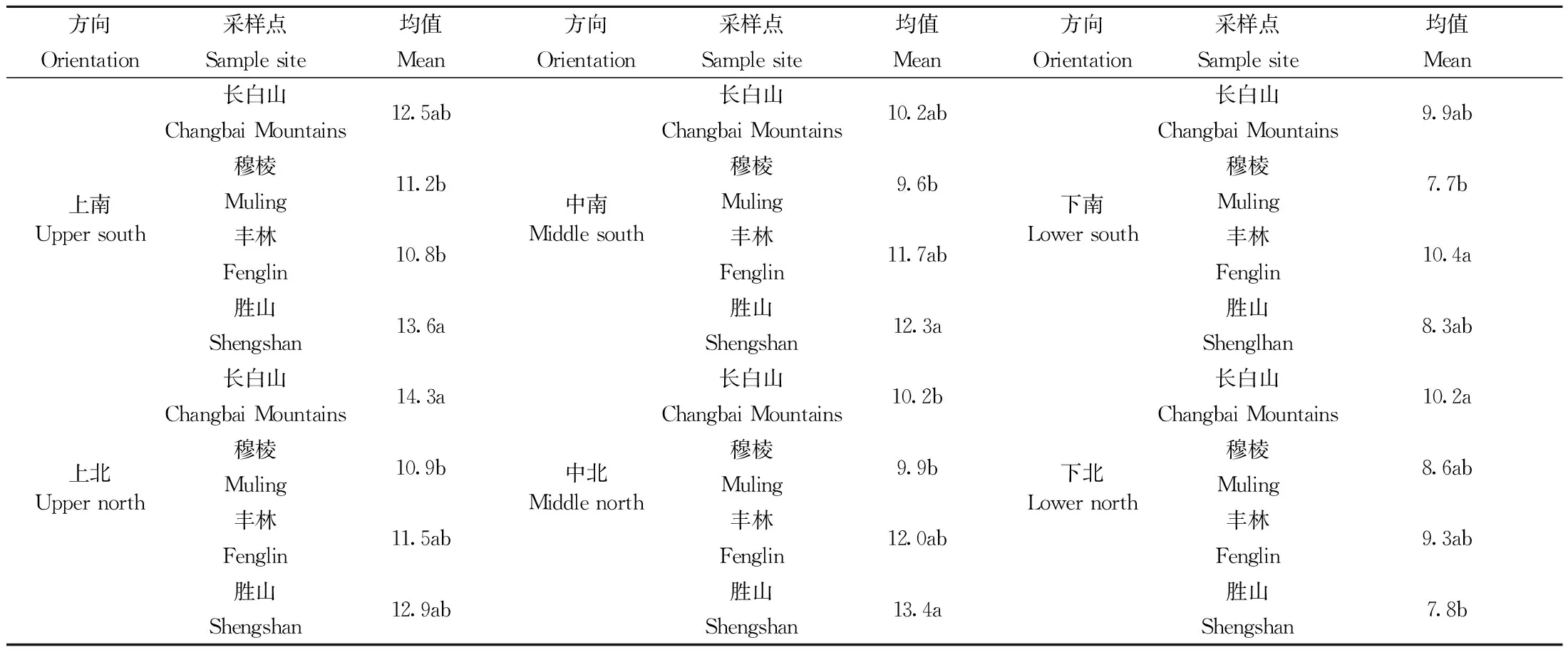
表4 红松枝横截面积在不同纬度、不同方向的差异Tab.4 Difference of twig cross-sectional areas of P. koraiensis in different latitudes and orientations
2.2 红松枝叶性状关系的纬度差异
不同纬度红松枝截面积与出叶强度均呈负相关,与叶面积呈正相关,但在不同纬度间,这些关系的斜率存在差异(P<0.05) (图2)。如在胜山(纬度最高),枝叶性状关系的斜率最小,枝横截面积与单叶面积的斜率为0.534 7、与总叶面积的斜率为1.352、与出叶强度的斜率为-1.661。

图2 红松枝叶性状关系的纬度差异性Fig. 2 Difference in relationship between twig and leaf traits of P. koraiensis along latitude低: 低纬度采样点(棕色); 中低: 中低纬度采样点(蓝色); 中高: 中高纬度采样点(绿色); 高: 高纬度采样点(红色)。不同颜色的公式代表不同采样点。P值表示枝叶性状的相关程度。P < 0.05代表性状显著相关; P> 0.05代表性状不显著相关,无回归关系线。P值表示不同纬度差异程度,P < 0.05表示不同纬度斜率具有显著差异。Low: low latitude sampling site (brown);mid low: Middle low latitude sampling site (blue); Mid high: middle high latitude sampling site (green); High: high latitude sampling site (red). The different colors of formulas represent different sampling sites.The P value in the figure indicates the degree of relationship in trait-trait, P< 0.05 indicates that there is significant relationship in trait-trait. If P> 0.05 indicates that there is no significant relationship in trait-trait, sono regression line. The P value in the figure indicates the degree of difference between different latitudes,P< 0.05 indicates that the slope is significantly different at different latitudes.
在4个采样点,枝横截面积、单叶面积和总叶面积之间呈异速生长关系,即其斜率与1存在差异,随着纬度减小,枝横截面积与出叶强度的斜率增大,并与-1之间差异显著,表现为异速生长关系; 在胜山采样点,单叶面积与出叶强度呈显著负相关关系,斜率为-0.332 0,与-1差异显著,为异速生长关系(图2)。
3 讨论
3.1 红松枝叶性状的纬度差异
自然选择更倾向于选择小叶子和高出叶强度红松叶面积、出叶强度和枝横截面积在不同纬度差异显著(表3),可能源于其选择的高出叶强度(Kleimanetal., 2007; Yangetal., 2008; Milla, 2009)。适度提高出叶强度可大大减少叶片受损的可能,而高出叶强度也将大幅度降低食草昆虫产生的危害(Molesetal., 2000; Kleimanetal., 2007)。在较低纬度的长白山,单叶面积相对较小,可能是低纬度温度相对较高所致(表1),其选择小叶片可以相对减少蒸腾导致的失水(Gates, 1980; Ackerlyetal., 2002; Osadaetal., 2015)。纬度最高区域,温度最低,总叶面积最小,可能是低温抑制植物生长并降低其能量利用能力,导致光合作用的反馈抑制(Huangetal., 2014; Lietal., 2018),从而使植株对总叶面积的投入也相应减少。长白山、胜山和丰林的枝横截面积无显著差异,说明在进化过程中,红松对环境的适应策略是趋向于改变资源获取器官的投资(Hilbert, 1990; Dewar, 1993)。
枝叶性状变化的枝水平解释率较高,既可能源于植株个体发育的差异,也可能源于分层采样方式。本研究选择对6个方向不同冠层采样,是为了更好表征植株个体内部的性状差异。在穆棱采样点,枝横截面积在各方向(表4)与总体趋势(表3)一致,表明在纬度差异较大的情况下,全向采样可以反映各方向纬度差异; 另3个采样点与整体趋势不一致,表明不同采样位置对试验结果有一定影响。为了突显主要问题,应该针对不同研究目的选择不同采样方法,针对相同研究目的选择相同采样方法。此外,对单叶面积变化的纬度解释率最高为58%,超过枝及个体水平(图1)。不同性状间的纬度解释率存在显著差异,基于此,本研究进一步探讨了纬度对红松枝叶性状关系的影响,以期更深入了解红松枝叶性状在不同纬度间的权衡策略。
3.2 红松枝叶性状关系的纬度差异
不同红松枝横截面积与总叶面积、单叶面积均呈显著正相关,与出叶强度呈显著负相关。单叶面积与枝横截面积的关系符合“Corner’s rules”,即较粗枝上对应较大叶面积(White, 1983a; 1983b)。另外,根据“管道模型”理论,枝上叶表面积与其导管横截面积呈正比(Shinozakietal., 1964; Niklas, 1992),且枝横截面积与导管直径呈正比(Fanetal., 2017),因而,枝上总叶面积与枝横截面积呈正相关。一般情况下,枝叶大小呈正相关,与叶数量呈负相关,叶数量、叶面积、枝横截面积三者之间的权衡导致出叶强度与枝横截面积呈负相关(Kleimanetal., 2007; Yangetal., 2008; Milla, 2009)。在高纬度的胜山,其出叶强度与单叶面积呈负异速生长关系,可能是因为植物体内总生物量有限,导致不能兼得高出叶强度和大单叶面积(Kleimanetal., 2007; Huangetal., 2016)。 另外,单叶面积与枝横截面积呈正相关,而大枝更倾向于生长较少叶片(Yagi, 2004;杨冬梅等, 2012),进而使出叶强度与单叶面积呈负相关。
不同纬度红松单叶面积与枝横截面积的异速生长指数小于1,说明单叶面积增长速度小于枝横截面积(图2)。采样点为阔叶红松林分布区,地处东北,属典型温带季风气候,由于水分条件即海陆分布限制,降水量在夏季较多且时间较集中; 该气候环境下,叶片更易受到外力压折,保留较小叶片或减缓叶片增长速率可更好地适应环境(Olsonetal., 2009; Yanetal., 2013)。红松为常绿针叶树,在寒冷气候下会相对增加茎干的投入,以缓解冰冻引起的栓塞和干旱引起的气穴现象(Cavender-Baresetal., 2010; Gleasonetal., 2016)。此外,植物面对不同生境会采取不同的适应策略。如大叶不仅会增加光拦截和碳收益,而且因其叶边缘的热交换能力较弱,更适宜在较凉的环境中生存(Gates, 1980; Ackerlyetal., 2002; Yangetal., 2008); 而小叶的热交换能力相对较高,更适合在强光、高温、干旱和低营养的环境中生存。不同纬度红松总叶面积与枝横截面积的异速生长指数均大于1,表明在各采样点枝横截面积的增长速率均显著小于总叶面积(图2)。枝横截面积的增长速率大于单叶面积,但显著小于总叶面积,表明随着枝横截面积增加,植物更趋向于增加叶片数量以适应环境变化,尽管叶片数量增加将增大总投资成本,但其叶的多年生特性会在降低建构成本的同时增大总光合收益(Yangetal., 2010)。
出叶强度与单叶面积呈负相关的异速生长关系,且异速生长指数绝对值小于1,与李曼等(2017)研究结果一致。这说明“出叶强度优势”中的等速生长关系在种内水平可能并不适用,尤其在分析常绿针叶植物种内变异时(Kleimanetal., 2007)。
此外,在不同纬度,枝叶性状关系的斜率差异显著(图2)。采样点地处东北区域,决定植物生长的重要因子是温度,较高的温度将弥补低温生境容易造成的枝栓塞和低传播效率等问题(Cavender-Baresetal., 2010; Lietal., 2017),枝横截面积增大,其能量运输效率及水力运输显著提升,植物对叶片投入增加,将利于提高碳获取和营养利用效率(Hilbert, 1990; Dewar, 1993)。因此,高纬度地区温度较低,其单叶面积随着枝横截面积的增加速度显著低于低纬度地区(图2)。生境压力下,植物将相应增加对茎的投资,以减轻养分和水传输的压力,与Xiang等(2009)的发现一致。同时,降水差异直接改变了环境的湿润度。如在降水量低的地区,大叶子的强蒸腾作用会增加植物水分流失,因此植物更倾向于着生小叶片(Xiangetal., 2009;Osadaetal., 2015); 当降水较高且水量充足时,植物更趋向着生大叶片。在不同生境下,同种植物可能会采取一系列的适应性策略。因此,在年均温度较高的低纬度地区,单叶面积普遍较小(表1); 而降雨量相对较高,导致单叶面积随枝横截面积的增加速度高于高纬度地区(图2)。随着纬度降低,枝横截面积与总叶面积的异速生长指数呈先增加后减少的趋势,这说明一定范围内温度升高减缓了气候胁迫,在相对适宜环境中,植物会优先增大总叶面积;达到某个阈值后,过高温度将会限制植物在叶片上的投资(Cobleetal., 2015)。随着纬度降低,枝横截面积与出叶强度的异速生长指数增加,这说明在枝横截面积增加到一定值时,纬度越低,出叶强度下降越多。为了最大化资源收益,植物会在单位枝上分配数量较少的大叶片或较多的小叶片。在低纬度的长白山区域,红松的单叶面积较小(表3),因此其选择增大出叶强度以提高光捕获能力和利用效率(Molesetal., 2000)。
4 结论
枝叶性状间关系及单性状的变化,均反映了植物调整自身组织以应对不同生境响应策略的特性。本研究中,红松枝叶性状关系在不同纬度存在差异,其斜率在高纬度地区相对较小,表明植物会采取不同资源分配策略以适应环境变化,且枝叶协同适应环境变化。通过分析红松枝叶性状及其间的异速生长差异,可更好阐述其种内变异,为准确预测红松的生境响应提供参考。
