云南地方鸡种质资源现状与分子研究进展
2021-07-14王麒淋李广玉郭爱伟周杰珑陈粉粉李青青
王麒淋 李广玉 郭爱伟 周杰珑 陈粉粉 李青青
摘要:云南多样化的地理气候条件,形成了丰富的地方鸡种资源,有13个鸡种被列入《中国畜禽遗传资源志》,品质和数量均居全国之首。这些独特地方鸡种是改良培育鸡品种不可缺少的原始遗传素材和宝贵的基因储存。但由于种种原因,云南多种地方鸡种群体数量下降或面临消失,种质资源亟待维系,优良性状开发极大受限。在查阅大量相关研究报道的基础上,综述了云南省种质资源现状与分子研究进展,以期为地方核心种质资源的保护提供科学依据,为选育、复壮和扩群培育出更具独特风味的云南地方特有资源品种提供理论指导。
关键词:云南;地方鸡种;种质资源;分子研究进展
中图分类号:S831.2 文献标识码:A 文章编号:1007-273X(2021)04-0014-06
云南多样化的地理、生态、气候条件,众多的民族及不同的生活习惯,加之长期以来的驯养和培育,形成了丰富多彩的地方鸡种遗传资源,不仅物种多样、类群齐全,而且种质特性各异,在我国居各省(区)之首,其中13个鸡种被列入《中国畜禽遗传资源志(家禽志)》[1],云南六大名鸡驰名中外(大围山微型鸡、无量山乌骨鸡、武定鸡、茶花鸡、瓢鸡和盐津乌骨鸡)。这些独有地方家禽品种能适应当地生态环境和饲养条件,具有就巢性强、耐粗饲、肉质营养丰富等优点,更重要的是这些家禽品种具有很强的抗病性,如茶花鸡[2]和独龙鸡[3]等。这些独特的优良性状是众多民族在相对封闭的地域环境下长期驯养而成,是改良培育鸡品种不可缺少的原始遗传素材和宝贵的基因库,如茶花鸡一直被认为对我国家鸡遗传多样性的品种贡献率最高[4]。此外,这些独特的优良性状也是云南省禽业可持续发展的宝贵资源,是开发云南省特色禽产品,提高其禽业竞争力的潜在优势。
1 云南地方鸡种质资源现状
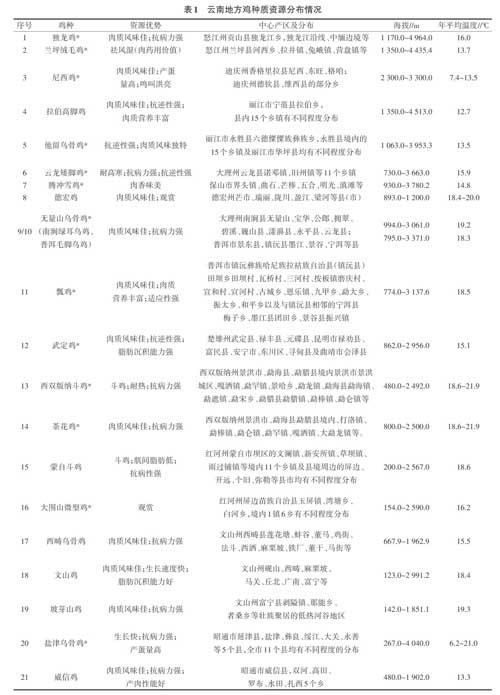
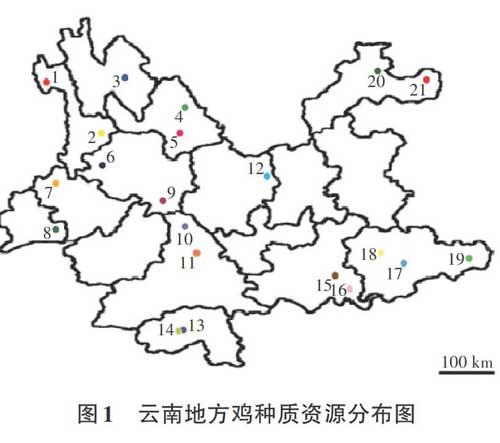
目前云南许多地方家禽品种养殖主要是靠农户自繁自养,没有统一的良种繁育体系,缺乏系统的选种选育和提纯复壮,鸡群多处于小区域的封闭状态,血缘近亲化严重[5]。再加上近年来,为满足人民群众对禽产品的需求和随着交通条件的改善,云南省相继引进了大量的外来高产品种,对低产地方品种进行改良,使禽业生产水平大幅度提高。但这些高产专门化品种(品系)在大幅提高畜牧业产量和效益的同时,也由于连续定向选择,使得品种(系)内、品种(系)间的遗传变异越来越窄,导致遗传资源锐减,同时也使某些地方品种逐渐被培育品种或杂交种所取代,致使具有丰富多样性的地方品种群体数量下降或消失,品种资源流失严重。据调查报道有些纯种的地方鸡种,如独龙鸡,目前仅存300只左右,濒临灭绝[6]。如果再不采取有效的保护措施,云南许多独有地方家禽品种的许多优良基因(如抗病基因)和经济性状(如肉品质)将会消失。因此,对其进行研究和保护迫在眉睫[3,7](表 1、图1)。
2 云南地方鸡种分子研究进展
2.1 线粒体基因
作为“能量工厂”和有氧呼吸主场所的细胞器,线粒体在细胞中发挥着重要作用[8]。线粒体DNA(Mitochondrial DNA, mtDNA)具有进化快、母系遗传和极少发生重组等特点,一直被认为是进行家鸡遗传多样性研究的首选工具[9]。
胡文平等早在1999年从血液蛋白、细胞遗传和mtDNA等方面证明红色原鸡为家鸡的祖先,并提出云南可能是中国家鸡的起源中心之一,采用线粒体hypervariable segment I(HVS-I)母系遗传标记的研究表明,家鸡母系世系可划分为9个支系,推測起源于包括:中国云南、南亚、东南亚及周边广大区域[10]。基于mtDNA全基因组序列构建系统发育树,进一步揭示在南亚和中国云南及其邻近地区发生过驯化事件,表明云南及其周边地区是家鸡中部分世系的重要起源地[11]。除了利用mtDNA探究推测家鸡的系统起源以外,还基于线粒体全基因组测序,快速探究不同鸡种遗传多样性等信息,童晓梅等[12]获得了藏鸡(Tibetan chicken)线粒体全基因组序列,其全长16 783 bp,共有13个蛋白质编码基因、2个rRNA基因、22个tRNA基因和1个D-loop。Yan等[13]获得大围山微型鸡线粒体全基因组序列,其全长16 785 bp,共包含37个基因(2个rRNA基因,13个编码蛋白质基因,22个tRNA基因)和1个D-loop。
线粒体中的控制区(Control region),也称D-环(Displacement loop,D-loop),是线粒体基因组中进化速率最高、最具多态的区域,广泛运用于家禽遗传学研究[9,14,15]。
控制区全序列方面,Jia等[16]扩增了大围山微型鸡线粒体D-loop区全序列,结合GenBank上5只红色原鸡(Gallus gallus)对应序列进行分析,表明大围山微型鸡D-loop区全长为1 231或1 232 bp,差别为859处是否缺失了碱基C。线粒体D-loop区全序列也被用来探究大围山微型鸡[17-19]、茶花鸡[17,20]、云龙矮脚鸡[19]和拉伯高脚鸡[19]的遗传多样性和起源进化。结果表明,这些地方鸡均具有丰富的遗传多样性且有多个母系起源,但红色原鸡滇南亚种在鸡种形成过程中贡献最大。Zhang等[21]测定了239个采自云南迪庆等地的藏鸡和283个四川盆地家鸡的线粒体控制区片段,结合下载的1 091个GenBank中已报道序列,对藏鸡[22]的起源和高海拔低氧适应性进化进行了研究。
控制区部分序列方面也有不少研究。独龙鸡仅分布于我国云南省贡山县,是极其濒危的地方特有鸡种,Li等[23]对59羽独龙鸡mtDNA的D-loop区进行分析,揭示了独龙鸡的单一母系血统。包文斌等[24]基于mtDNA的D-loop区560 bp序列,研究了14个中国地方鸡种和红色原鸡的遗传多态性与系统进化。结果显示,每个群体表现出较高水平的遗传多态性;群体间表现出显著的遗传分化;固始鸡和仙居鸡起源于泰国红色原鸡亚种(Gallus gallus gallus),而茶花鸡和藏鸡起源于中国红色原鸡亚种(Gallus gallus spadiceus)。刘丽仙等[25]对云南8个地方鸡群体mtDNA的D-loop区遗传多样性及系统进化进行了分析。其他研究者对云南地方鸡种的研究还包括他留乌骨鸡、兰坪绒毛鸡和武定鸡[26]、普洱毛脚乌鸡与南涧绿耳乌鸡[27]、华坪乌骨鸡[28]、瓢鸡[29]、西畴乌骨鸡[30]等。
2.2 微卫星标记
微卫星DNA(Microsatellite),又称短串联重复序列(Short tandem repeats,STR)或简单重复序列(Simple sequence repeats,SSR),是以少数几个核苷酸(一般2~6个)为单位多次串联重复的DNA序列,广泛均匀分布于真核基因组中[31]。微卫星DNA具有多态性高、共显性遗传、操作简单、检测容易、适合进行自动化分析和引物保守性(Cross-priming strategy)[32, 33]等诸多优势,也是应用于禽类遗传多样性、群体结构与分化及亲缘关系构建的另一个重要分子标记。
尼西鸡和茶花鸡是分布仅限于香格里拉和西双版纳的2种云南省特有地方鸡种,具有多种优良性状(尤其是抗逆性),均已被列入国家畜禽遗传资源目录[4,34,35]。叶朗惠等[34,35]利用家鸡24条染色体上的33个微卫星标记对尼西鸡和茶花鸡群体遗传变异进行了检测分析,等位基因数量、群体平均表观杂合度、期望杂合度及平均多态信息含量等数据均表明尼西鸡和茶花鸡群体内存在丰富的遗传多样性。利用同样的微卫星标记,对多种云南地方鸡种进行了研究,包括腾冲雪鸡[36]、文山乌骨鸡[37]、屏边大围山微型鸡[38]和孔雀绿雉鸡[39]等。
Huo等[40]运用28个微卫星DNA标记对云南7个鸡种(包括红色原鸡)共340只鸡样本进行了分型。结果表明,在已鉴定的342个等位基因中有121个(35.4%)具有品种特异性,其中红色原鸡等位基因数目最多,为23个;群体内杂合度较高,平均无偏HE值为0.663,高于其他群体。利用18对微卫星引物分析了4个地方特色鸡种(藏鸡、勐腊茶花鸡、贡山土鸡和武定鸡)的遗传多样性,系统聚类树显示武定鸡和贡山土鸡关系最近,首先聚为一类;其次与勐腊茶花鸡聚为一类,形成云南支,最后与藏鸡聚为一类,完全符合群体间的地理格局[41]。曲鲁江等[42]利用分布于13条染色体上的27个微卫星标记,探讨了分布于中国各地的78个地方鸡品种遗传多样性,并通过Nei氏遗传距离和邻接法将中国地方鸡种进行聚类分析。结果显示,中国的地方鸡品种具有广泛的遗传多样性,遗传关系与其地理分布关系密切,大体上可分为6类(西北型、华南型、华东型、西南I型、西南型Ⅱ型和华中型)。该研究中涉及了6个云南地方鸡种,分子聚类树显示西双版纳斗鸡、茶花雞和腾冲雪鸡都属于华南型;武定鸡和云龙矮脚鸡属于西南型Ⅱ型;盐津乌骨鸡属于其他型。李慧芳等[43]通过选用30个多态性较好的微卫星标记,检测了云南省武定鸡、茶花鸡、云龙矮脚鸡、盐津乌骨鸡、腾冲雪鸡和西双版纳斗鸡共6个地方鸡品种的遗传多样性。结果显示,武定鸡杂合度最低,腾冲雪鸡杂合度最高;6个鸡品种被聚为3类:第1类为云龙矮脚鸡、盐津乌骨鸡和武定鸡,第2类为腾冲雪鸡和西双版纳斗鸡,茶花鸡独自为最原始的一类,这与6个鸡种的来源、分化、选育历史及地理分布一致。
包文斌等[44]利用29个微卫星DNA标记对来自中国的红原鸡(Gallus gallusspadiceus)和来自泰国的红原鸡(Gallus gallus gallus)进行了比较分析,认为中国家鸡为独立起源。汤青萍等[45,46]用27对微卫星引物对中国5种斗鸡和12个地方乌骨鸡品种进行遗传多样性分析。其他研究还包括盐津乌骨鸡[47]、版纳斗鸡[48]、茶花鸡[49-52]、藏鸡[49-52]和武定鸡[53]。
2.3 功能基因研究
家鸡作为人类饲养范围最广泛的家禽[54],随着饲养规模化和集约化发展,其抗病性和生产性能相关的功能基因备受关注[55]。周勇等[56]对危害养鸡业发展的三大疫病之一的马立克氏病(MD)抗性相关基因进行了综述。王广龙等[57]通过了解武定鸡群白血病病毒受体TVB基因型分布情况,为下一步研究禽白血病病毒(Avian leukosis virus,ALV)——B、D、E亚群禽白血病抗病育种奠定了基础。段博芳等[58]通过采用Real-time荧光定量PCR的方法获得云南6个特色鸡种(大围山微型鸡、武定鸡、茶花鸡、无量山乌骨鸡、盐津乌骨鸡和瓢鸡)不同日龄禽流感Mx1基因的表达差异情况,可为今后制订抗病育种方案提供参考依据。新城疫(Newcastle disease)是对家鸡养殖危害最大的疾病之一,具有传播快、致死率高的特性[59],近期研究表明,鸡近交系中eIF2基因家族的差异表达有助于抵抗新城疫[60]。
MC1R基因是控制动物黑色素合成的重要基因,也是鸡黑色素性状的主效基因或是与鸡控制黑色素性状的主效基因连锁。王建等[61]对白羽肉鸡和黑羽乌骨鸡的资源家系及由该亲本建立的资源家系群体的MC1R基因进行PCR-SSCP分析,分析其性状与基因型的相关性,不仅为合理开发利用乌骨鸡这一宝贵的家禽资源提供了理论依据,也为进一步杂交利用提供了可靠的分子检测手段。张学余等[62]利用随机扩增多态性DNA(Random amplified polymorphic DNA,RAPD)技术对泰和鸡、固始鸡、萧山鸡、狼山鸡鸡种性染色体上含有Id和id基因进行了正反交试验,测定了其肤色的基因型,探明肤色伴性基因在产生的RAPD共显性条带的差异。
脂肪分化相关蛋白(ADFP)、脂肪酸转运蛋白1(FATP1)和载脂蛋白B(APOB)在确定肌内脂肪和肉质中起着重要作用[63]。Li等[63]检测了大围山微型鸡、盐津乌骨鸡、茶花鸡、武定鸡、无量山乌骨鸡和瓢鸡不同发育阶段的不同组织中ADFP基因、FATP1基因和APOB基因的表达和脂肪性状相关的情况,为更加合理开发利用宝贵的地方鸡种资源提供了一定的理论依据。Zhang等[64]研究了FGF1和FGF10基因在藏鸡5个发育阶段的表达情况,阐明了这2个基因在藏鸡脂肪细胞发育和肌肉脂肪沉积的功能。GnRHI和GnRHII基因在鸡排卵和产蛋中起着关键作用,研究表明GnRHI和GnRHII基因具有多态性,对白来航蛋鸡体重、产蛋量和蛋品质性状有显著影响[65]。SH3RF2基因位于家鸡体重相关的数量性状位点,它的缺失与家鸡体重成正相关[66,67]。
豆腾飞等[68]研究表明,GS基因和ADSL基因[69]均是影响家禽肉品质的重要候选基因,TNF-α基因[70]对家禽的骨强度有重要影响。李美荃等[71]对普洱瓢鸡、腾冲雪鸡与三黄鸡8个组织进行实时荧光定量,探讨了抗氧化基因GRX1(谷氧还蛋白)和TRX1(硫氧还蛋白)在不同鸡种、组织间的差异表达,表明这可能是影响本地鸡种与培育鸡种抗氧化能力的重要途径之一。王春光等[72]基于云南4个地方鸡种PRL外显子5基因的多态性,证明其A基因可能是影响繁殖性状的优势基因。
3 结语
云南多元化的地理、生态和气候条件,加上长期以来经过广大劳动人民的精心选育,形成了丰富的地方鸡种资源。本文主要从分子水平上综述了云南部分地方鸡种种群遗传结构和分化程度;阐明了一些种群间亲缘关系和起源进化信息;探讨了与家鸡生产性能紧密关联的多种基因。这些结果可为挖掘云南地方优良性状相关基因和品种改良培育提供线索,同时为进一步制定云南特色禽类种质资源的保护和利用方案提供重要依据。
参考文献:
[1] 苗永旺, 葛长荣. 云南地方鸡资源的发掘、保护与利用[J]. 中国家禽, 2019, 41(23): 1-8.
[2] 刘建平, 和俊权, 波 内. 云南省西双版纳茶花鸡种质资源保护探索[J]. 当代畜牧,2016(8Z): 19-20.
[3] 文生萍, 胡骁飞, 岳炳辉, 等. 独龙鸡肉品质及血液生化指标的测定[J]. 黑龙江畜牧兽医, 2017, 534(18): 69-71.
[4] 李慧芳, 朱云芬, 韩 威, 等. 中国地方鸡种遗传多样性和品种贡献率分析[J]. 家畜生态学报, 2009, 30(5): 14-15.
[5] 卓 根. 特种养殖 您有风险意识吗?[J]. 猪业观察, 2007 (19): 43.
[6] 文生萍, 胡骁飞, 高明艳, 等. 云南独龙鸡体尺性状和屠宰性能测定及相关性分析[J]. 浙江农业科学, 2017, 58(1): 147-149.
[7] 云南省畜禽遗传资源委员会. 云南省畜禽遗传资源志[M]. 昆明:云南科技出版社, 2014.
[8] SHARMA P, SAMPATH H. Mitochondrial DNA integrity:Role in health and disease[J]. Cells, 2019, 8(2): 100.
[9] LAN D, HU Y, ZHU Q, et al. Mitochondrial DNA study in domestic chicken[J]. Mitochondrial DNA Part A, 2017, 28(1): 25-29.
[10] LIU Y P, WU G S, YAO Y G, et al. Multiple maternal origins of chickens:Out of the Asian jungles[J]. Molecular phylogenetics and evolution, 2006, 38(1): 12-19.
[11] MIAO Y W, PENG M S, WU G S, et al. Chicken domestication: An updated perspective based on mitochondrial genomes[J]. Heredity (Edinb), 2013, 110(3): 277-282.
[12] 童曉梅, 梁 羽, 王 威, 等. 藏鸡线粒体全基因组序列的测定和分析[J]. 遗传, 2006, 28(7): 769-777.
[13] YAN M L, DING S P, YE S H, et al. The complete mitochondrial genome sequence of the Daweishan Mini chicken[J]. Mitochondrial DNA Part A, 2016, 27(1): 138-139.
[14] NISAR A, WAHEED A, KHAN S, et al. Population structure, genetic diversity and phylogenetic analysis of different rural and commercial chickens of Pakistan using complete sequence of mtDNA D-loop[J]. Mitochondrial DNA Part A, 2019, 30(2): 273-280.
[15] NUNOME M, KINOSHITA K, ISHISHITA S, et al. Genetic diversity of 21 experimental chicken lines with diverse origins and genetic backgrounds[J]. Experimental animals, 2018,68(2): 177-193.
[16] JIA X X, TANG X J, LU J X, et al. The investigation of genetic diversity and evolution of Daweishan Mini chicken based on the complete mitochondrial (mt)DNA D-loop region sequence[J]. Mitochondrial DNA, 2016, 27(4): 3001-3004.
[17] 陆俊贤, 贾晓旭, 唐修君, 等. 2个云南原始鸡种遗传多样性及其与红色原鸡的亲缘关系[J]. 浙江大学学报(农业与生命科学版), 2016, 42(3): 385-390.
[18] 贾晓旭, 唐修君, 陆俊贤, 等. 基于线粒体DNAD-loop区全序列分析大围山微型鸡遗传多样性及其起源进化关系的研究[J]. 南京农业大学学报, 2015(4): 656-660.
[19] 欧阳依娜, 阮 谦, 钱林东, 等. 大围山微型鸡、云龙矮脚鸡和拉伯高脚鸡mtDNA D-loop遗传多样性分析[J]. 云南农业大学学报(自然科学版), 2017 (1): 70-77.
[20] 范梅华, 唐修君, 贾晓旭, 等. 基于线粒体D-loop区分析茶花鸡遗传多样性和起源进化[J]. 中国家禽, 2019, 41(20): 8-11.
[21] ZHANG L, ZHANG P, LI Q Q, et al. Genetic evidence from mitochondrial DNA corroborates the origin of Tibetan chickens[J]. PLoS ONE, 2017, 12(2): e0172945.
[22] WANG M S, LI Y, PENG M S, et al. Genomic analyses reveal potential independent adaptation to high altitude in Tibetan chickens[J]. Molecular biology and evolution, 2015, 32(7): 1880-1889.
[23] LI Q, ZHANG P, LI M, et al. Genetic diversity and relationship of Dulong chickens using mitochondrial DNA control region[J]. Mitochondrial DNA Part B, 2020, 5(1): 275-280.
[24] 包文斌, 陈国宏, 李碧春, 等. 红色原鸡和中国家鸡遗传多样性及亲缘关系[J]. 中国科学(C辑:生命科学), 2008(1): 43-51.
[25] 刘丽仙, 戚晨阳, 佟荟全, 等. 云南8个地方鸡群体mtDNA D-loop区遗传多样性及系统进化分析[A]. 遗传多样性:前沿与挑战——中国的遗传学研究(2013-2015)——2015中国遗传学会大会论文摘要汇编[C]. 北京:中国遗传学会,2015.
[26] 苗永旺, 孙利民, 王素芳, 等. 云南3个地方鸡群体mtDNA D-loop遗传多样性分析[J]. 云南农业大学学报(自然科学版), 2015, 30(3): 395-401.
[27] 苗永旺, 孙利民, 童晶晶. 普洱毛脚乌鸡与南涧绿耳乌鸡线粒体DNA母系遗传分析[J]. 家畜生态学报, 2013, 34(7): 10-14.
[28] 刘丽仙, 苗永旺, 孙利民, 等. 华坪乌骨鸡mtDNAD-loop遗传多样性分析[J]. 畜牧与兽医, 2012, 44(10): 30-33.
[29] 贡潘偏抽, 刘丽仙, 李大林, 等. 基于线粒体DNA控制区(mtDNA D-loop)序列分析瓢鸡的遗传多样性[J]. 云南农业大学学报(自然科学版), 2011, 26(2): 211-214.
[30] 黄道平, 李大林, 袁 峰, 等. 西畴乌骨鸡mtDNA D-loop区遗传多样性分析[J]. 云南农业大学学报(自然科学版), 2010, 25(3): 373-376.
[31] 易 冰, 李卓雅, 孟锦绣, 等. 微卫星DNA在寄生虫学研究中的应用[J]. 中国人兽共患病学报, 2003, 19(2): 85-87.
[32] MOORE S, SARGEANT L, KING T, et al. The conservation of dinucleotide microsatellites among mammalian genomes allows the use of heterologous PCR primer pairs in closely related species[J]. Genomics, 1991, 10(3): 654-660.
[33] SCHLOTTER?ER C, AMOS B, TAUTZ D. Conservation of polymorphic simple sequence loci in cetacean species[J]. Nature, 1991, 354(6348): 63-65.
[34] 葉朗惠, 苗永旺, 霍金龙, 等. 茶花鸡群体遗传多样性[J]. 动物学杂志, 2006, 41(2): 37-42.
[35] 叶朗惠, 霍金龙, 苗永旺, 等. 尼西鸡遗传多样性微卫星标记分析[J]. 动物学研究, 2006, 27(1): 68-74.
[36] 苗永旺, 陈 涛, 霍金龙, 等. 利用微卫星标记分析腾冲雪鸡的遗传多样性[J]. 西南农业学报, 2008, 21(5): 1431-1433.
[37] 苗永旺, 叶朗惠, 陈 涛, 等. 文山山地乌骨鸡微卫星DNA多态性分析[J]. 云南农业大学学报(自然科学版), 2009, 24(1): 63-66.
[38] 贾俊静, 文生萍, 熊保良, 等. 云南屏边大围山微型鸡遗传多样性[J]. 云南农业大学学报(自然科学版), 2009, 24(5): 654-660.
[39] 高姗姗, 付 琳, 袁 峰, 等. 雉鸡微卫星DNA标记筛选及孔雀绿雉鸡群体遗传变异分析[J]. 云南农业大学学报(自然科学版), 2014, 29(5): 677-683.
[40] HUO J, WU G, CHEN T, et al. Genetic diversity of local Yunnan chicken breeds and their relationships with Red Junglefowl[J]. Genetics and molecular research, 2014, 13(2): 3371-3383.
[41] 杜 宏, 黄燚娜, 关伟军, 等. 应用微卫星标记分析云南、西藏4个地方特色鸡种的遗传多样性[J]. 甘肃农业大学学报, 2011, 46(4): 7-12.
[42] 曲鲁江, 李显耀, 徐桂芳, 等. 利用微卫星标记分析中国地方鸡种的遗传多样性[J]. 中国科学:生命科学, 2006, 36(1): 17-26.
[43] 李慧芳, 陈宽维, 汤青萍, 等. 利用微卫星标记分析云南6个地方鸡品种的遗传多样性[J]. 江苏农业学报, 2006, 22(1): 33-37.
[44] 包文斌, 陈国宏, 吴信生, 等. 中国红原鸡和泰国红原鸡遗传多样性分析[J]. 遗传, 2007, 29(5): 587-592.
[45] 汤青萍, 陈宽维, 李慧芳, 等. 应用微卫星标记对12个中国地方乌骨鸡品种遗传多样性的研究[J]. 畜牧兽医学报, 2005, 36(8): 755-760.
[46] 汤青萍, 陈宽维, 李慧芳, 等. 中国斗鸡遗传分化的微卫星标记分析[J]. 西北农林科技大学学报 (自然科学版), 2005, 33(3): 19-23, 28.
[47] 陈 涛, 苗永旺, 霍金龙, 等. 盐津乌骨鸡微卫星DNA多态性研究[J]. 云南农业大学学报, 2007, 22(4): 543-546.
[48] 陈 涛, 霍金龙, 苗永旺, 等. 版纳斗鸡群体遗传多样性研究[J]. 云南农业大学学报, 2007, 22(3): 393-395, 400.
[49] 王得前, 陈国宏, 吴信生, 等. 运用微卫星技术分析中国地方鸡品种的亲缘关系[J]. 扬州大学学报(农业与生命科学版), 2003, 24(2): 1-6.
[50] 吴信生, 陈国宏, 王得前, 等. 利用微卫星技术分析中国部分地方鸡种的遗传结构[J]. 遗传学报, 2004, 31(1): 43-50.
[51] 高玉时, 杨 宁, 李慧芳, 等. 我国地方鸡品种保种群微卫星多态性分析与分子标记档案的建立[J]. 遗传, 2004, 26(6): 859-864.
[52] 陈国宏, 季从亮, 王敏强. 12个中国地方鸡种群体遗传结构及遗传多样性分析[J]. 畜牧兽医学报, 2006, 37(2): 105-111.
[53] 钱林东, 涛 陈, 霍金龙, 等. 武定鸡群体遗传变异的微卫星标记分析[J]. 云南农业大学学报, 2006, 21(5): 651-656.
[54] 梁素芸, 周正奎, 侯水生. 基于测序技术的畜禽基因组学研究进展[J]. 遗传, 2017, 39(4): 276-291.
[55] NAWAB A, AN L, WU J, et al. Chicken toll-like receptors and their significance in immune response and disease resistance[J].International reviews of immunology, 2019, 38(6): 284-306.
[56] 周 勇, 张 健, 杨斯涵, 等. 武定鸡抗马立克氏病相关基因的研究进展[J]. 中国畜牧兽医文摘, 2014, 30(5): 49-50.
[57] 王广龙, 王皓然, 王 勇, 等. 云南地区武定鸡白血病病毒受体TVB基因型的检测[J]. 中国家禽, 2017, 39(9): 11-15.
[58] 段博芳, 相德才, 吕 艳, 等. 基因在云南地方鸡种肝脏中的表达分析[J]. 中国家禽, 2017, 39(12): 54-56.
[59] 段雪梅. 鸡新城疫的诊断及防控[J]. 中兽医学杂志, 2018 (1): 33.
[60] DEL VESCO A P, KAISER M G, MONSON M S, et al. Genetic responses of inbred chicken lines illustrate importance of eIF2 family and immune-related genes in resistance to Newcastle disease virus[J]. Scientific reports, 2020, 10(1).DOI: 10.1038/s41598-020-63074-9.
[61] 王 建, 鐘发刚, 吴桐忠. MC1R基因多态性与其肤色、胫色表型相关性的分析[J]. 国外畜牧学-猪与禽, 2008, 28(2): 75-77.
[62] 张学余, 黄凡美, 周怀军, 等. RAPD技术对部分地方鸡种的肤色伴性遗传群体遗传结构的分析[J]. 江苏农业研究, 1999, 20(4): 3-5.
[63] LI J, ZHAO Z, XIANG D, et al. Expression of APOB, ADFP and FATP1 and their correlation with fat deposition in Yunnans top six famous chicken breeds[J]. British poultry science, 2018, 59(5): 494-505.
[64] ZHANG R, LI R, FENG Q, et al. Expression profiles and associations of FGF1 and FGF10 with intramuscular fat in Tibetan chicken[J]. British poultry science, 2018, 59(6): 613-617.
[65] BHATTACHARYA T K, CHATTERJEE R N, DANGE M, et al. Polymorphisms in GnRHI and GnRHII genes and their association with egg production and egg quality traits in chicken[J]. Britishpoultry science, 2019, 60(3): 187-194.
[66] RUBIN C J, ZODY M C, ERIKSSON J, et al. Whole-genome resequencing reveals loci under selection during chicken domestication[J]. Nature, 2010, 464(7288): 587-591.
[67] JING Z, WANG X, CHENG Y, et al. Detection of CNV in the SH3RF2 gene and its effects on growth and carcass traits in chickens[J]. BMC Genetics, 2020, 21(1): 22.
[68] 豆騰飞, 汪善荣, 刘丽仙. 武定鸡和艾维茵肉鸡GS基因表达量与肌糖原含量及pH值相关性研究[J]. 中国家禽, 2016, 38(8):17-20.
[69] 豆腾飞, 汪善荣, 佟荟全, 等. 武定鸡和大围山微型鸡肌苷酸含量及ADSL基因表达差异研究[J]. 中国家禽, 2017, 39(14):7-10.
[70] 豆腾飞, 荣 华, 张丽春, 等. 基因表达量对大围山微型鸡及武定鸡股骨强度的影响[J]. 中国家禽, 2015, 37(5):10-13.
[71] 李美荃, 宋 聪, 张春勇, 等. 云南普洱瓢鸡、腾冲雪鸡GRX1、TRX1基因差异性表达[J]. 家畜生态学报, 2015, 36(12):19-23.
[72] 王春光, 相德才, 叶绍辉. 云南4个地方鸡种PRL外显子5基因多态性研究[J]. 安徽农业科学, 2014, 42(17):5374-5376.
