“泉口冰雪柚”光合作用特性研究
2021-07-02陶爱群姜小文
陶爱群,姜小文
(湖南环境生物职业技术学院 园林学院,湖南 衡阳 421005)
柚(CitrusL.) 是芸香科(Rutaceae) 柑橘属(Citrinae)的一个种,果耐贮运,汁胞晶莹,风味佳,营养丰富,是热带、亚热带亚洲驰名的果品[1]。泉口冰雪柚是2008 年南方大范围冰灾后,从琯溪蜜柚果园冻后遗留琯溪蜜柚树中选育而来的柚类新品种。有关其它柚光合作用研究较多[2-9],但未见泉口冰雪柚光合特性的报道。笔者对泉口冰雪柚部分光合作用特性进行研究,不仅有助于了解其光合特性,而且也可以丰富柑橘光合作用研究的内容,同时为其适地引种和高光效栽培提供理论依据。
1 材料和方法
1.1 供试材料
供试材料为定植于湖南省衡阳县栏陇乡泉口村柚种植基地的5 a 生泉口冰雪柚。定植密度为2.5 m×3.0 m,常规管理。
1.2 研究方法
分别于2019 年7 月、9 月和11 月用Li-6400 测定净光合速率(Pn),气孔导度(Gs),胞间CO2浓度(Ci),蒸腾速率(Tr),叶片与空气间的水气压亏缺(VPD),气温(Tair),叶温(Tleaf),大气CO2浓度(Ca),样品室空气湿度(RH),光照强度(PAR)等。测定时分别选择外围有果春梢(psfw)、外围无果春梢(psfu)、内膛无果春梢(isp)、外围夏梢(psu)等枝条从上往下数第3~4 片完整功能叶,每次测3 株,每种类型叶测3 片。除光合作用的日变化外,其他测定均在当月日变化测量日的前后1~2 d 上午8∶00~11∶00 进行。
光合作用的日变化 在每月中下旬选择晴天测定日变化,7∶00~18∶00 每隔1 h 测定1 次。
光合作用的光响应曲线利用自动曲线测定功能,将红蓝光源设定光强梯度为0、400、800、1 200、1 600、2 000 μmol l/(m2s),测定叶片净光合速率。测定时样本室CO2浓度为400±1 μmol/mol,叶片温度30±1℃,相对湿度75±2%。
光合作用的CO2响应曲线(Pn-Ci)在0~1 000 μmol/mol范围内设定CO2浓度梯度(0、200、400、600、800、1 000、1 200 μmol/mol),测定净光合速率。测定时光强为1 000±2 μmol/(m2s),叶片温度设为30±1℃,相对湿度75±2%。
表观量子效率(AQY)在0~200 μmol/(m2s)低光强范围内,光强梯度设定为0、50、100、150、200 μmol/(m2s),测定Pn。测定时样本室CO2浓度400±1 μ mol/mol,叶 片 温 度30±1℃,相 对 湿 度75±2%。将Pn 和PAR 的成对值进行直线回归,回归方程的斜率即为表观量子效率。
羧化效率(CE)在0~250 μmol/mol范围内,将CO2浓度梯度设定为0、50、100、150、200、250 μmol/mol,测定Pn。测定时光强1 000±2 μmol/(m2s),叶温30±1℃,相对湿度75±2%,用Pn-CO2成对值进行回归,回归方程的斜率即羧化效率。
气孔限制值利用Berry 和Downton 的方法计算:
Ls=1-Ci/(Co-CCC)≌1-Ci/Co
1.3 数据分析
所有数据取平均值进行计算。补偿光强和补偿CO2浓度分别是表观量子效率和羧化效率回归方程截距与斜率的负数,饱和光强和饱和CO2浓度采用回归曲线法求出,所有数据和图表用Microsoft Excel 2010处理。
2 结果与分析
2.1 光合作用的日变化
田间泉口冰雪柚在不同月份、不同类型叶片、不同天气条件下日变化均不相同(图1-4)。
外围有果春梢叶片的净光合速率(Pn)日变化呈双峰曲线,出现“午休”现象,7 月、9 月、11 月(采果后)首峰分别出现在10:00、9:00、8:00;随后净光合速率出现不同程度的下降,7 月和9 月低谷值在14:00[6.62 μmol/(m2s)]和[1.49 μmol/(m2s)]、11 月在12:00[1.71 μmol/(m2s)];15:00、16:00、14:00 出现次峰,次峰值小于首峰值(图1)。气孔导度(Gs)和Pn变化趋势一致,叶片胞间CO2浓度(Ci)早晨最高,午休出现前与Gs 变化趋势相反,午休出现后Ci上升或变化不大(Gs 下降),上升幅度9 月最大、7 月最小,和午休低谷值大小相反,进一步分析光强(PAR)、叶片温度(Tleaf)和空气相对湿度(RH),发现7月时最大PAR[1 983 μmol/(m2s)]、最高Tleaf(38.01℃)和最小RH(34.74%)均高于9月[1 740μmol/(m2s)],34.73 ℃,16.35%和11月[1 638 μmol/(m2s)],31.62 ℃,15.63%,而Gs 的下降幅度却7 月小于9 月和11 月。同时,气孔限制值变化趋势(图4)也说明此时光合作用的的影响因素中,气孔因素占主导作用。综上分析说明7 月出现午休主要原因是高温和高光强引起气孔导度下降限制了CO2向叶肉细胞的扩散(气孔因素),高温低湿和高光强条件下的光合能力下降(非气孔因素)也有一定的影响,15∶00 时出现第二次峰值时气孔导度值并没有升高而胞间CO2浓度出现了下降证实了光合能力下降这一点;而9 月和11 月出现午休是高温低湿和高光强条件下气孔因素和非气孔因素共同作用的结果,第二峰值出现时气孔导度有所增大,而Ci有所下降,说明非气孔因素的影响减少。

图1 外围有果春梢晴天光合作用日变化Fig.1 Photosynthetic diurnal variation of psfw in sunny days
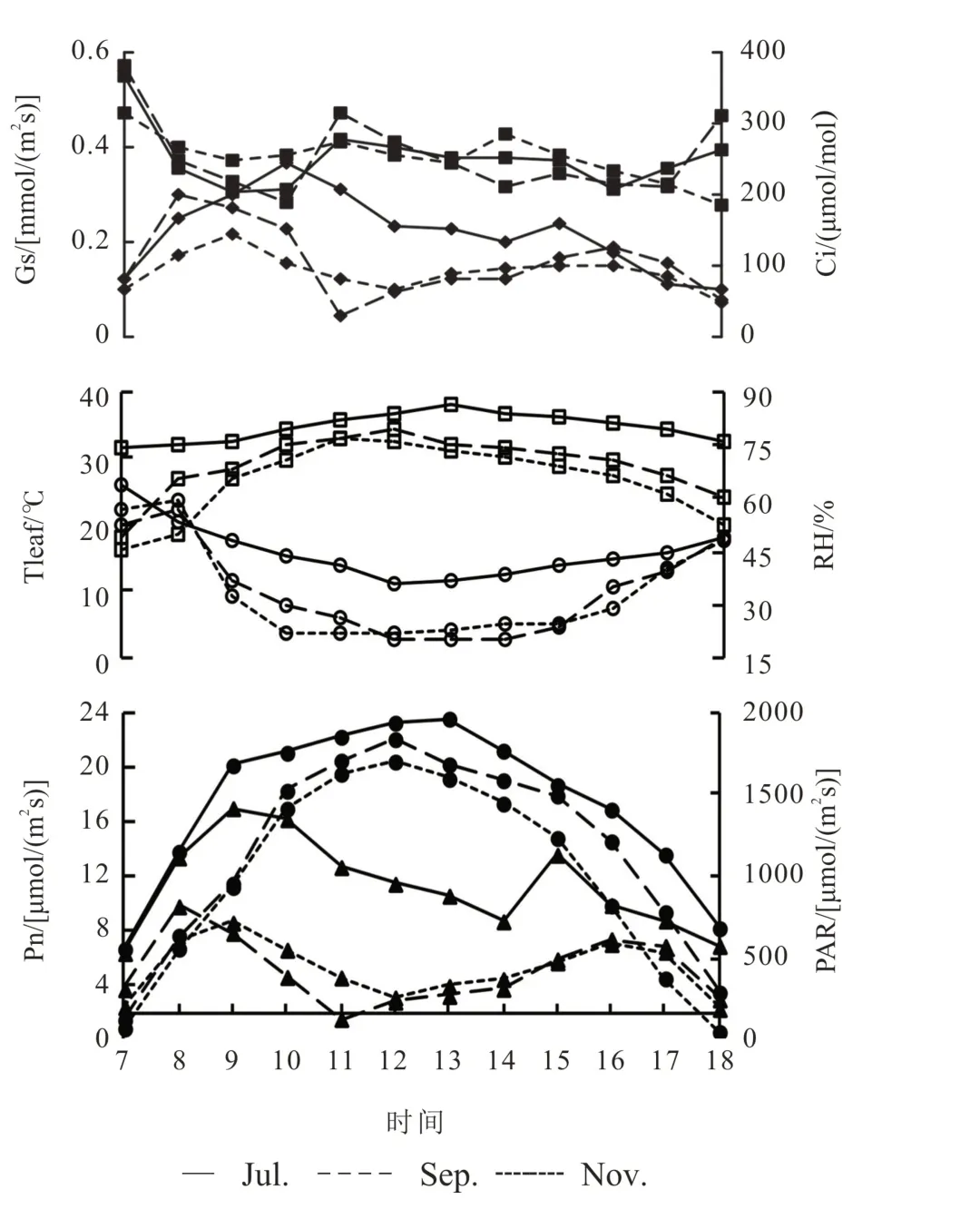
图2 外围无果春梢晴天光合作用日变化Fig.2 Photosynthetic diurnal variation of pspw in sunny days
外围无果春梢叶片晴天净光合速率(Pn)日变化和外围有果春梢相似,呈双峰曲线也出现午休现象,但峰值出现的时间和大小与外围有果春梢有差别(图1,2)。7 月、9 月、11 月首峰分别出现在11∶00[-0.38 μmol/(m2s)]、12∶00[1.19 μmol/(m2s)];9 月午休程度比外围有果春梢要深,净光合速率出现了负值;11月次峰比有果春梢明显,出现时间(16:00)晚于外围有果春梢。分析气孔导度(Gs)、叶片胞间CO2浓度(Ci)、光强(PAR)、叶片温度(Tleaf)和空气相对湿度(RH)等的变化趋势发现,7 月10:00气孔导度达到最大值时净光合速率已开始下降,说明此时午休的主要原因是非气孔因素,10:00 后气孔导度下降,胞间CO2浓度(Ci)不但没有下降反而略有升高,这说明光合午休是气孔因素(见图6)引起CO2供应不足和非气孔因素引起的光合能力下降的共同结果;9 月8∶00-10∶00 净光合速率随气孔导度和胞间CO2浓度下降而下降,主要是气孔因素的影响,随着温度的进一步升高和湿度的下降,非气孔因素的影响加大。气孔限制值的变化也表明了不同月份外围无果春梢叶片光合午休的影响因素因季节差异而变化。外围夏梢叶晴天净光合速率(Pn)和气孔导无果春梢叶片光合午休的影响因素因季节差异而变化。外围夏梢叶晴天净光合速率(Pn)和气孔导度(Gs)日进程呈双峰曲线,两者变化趋势较接近,说明净光合速率与气孔导度正相关,与春梢叶的结果一致。7 月出现午休时的胞间CO2浓度(Ci)不但没有下降反而略有升高,这说明光合午休不是气孔因素引起CO2供应不足,而是非气孔因素引起的光合能力下降,9 月和11 月出现午休时Ci 增加幅度较大,出现Pn低谷值[2.51 μmol/(m2s)和1.23 μmol/(m2s)]时相对湿度最低(16.27%和15.01%),这说明低湿等引起的非气孔因素导致了光合能力的下降,这是9 月和11 月光合午休的主要原因。气孔限制值变化(见图4)也应证了夏梢叶片7 月光合“午休”主要受气孔因素影响,9 月、11 月主要受非气孔因素影响。

图3 夏梢净光合作用日变化Fig.3 Photosynthetic diurnal variation of psu in sunny day
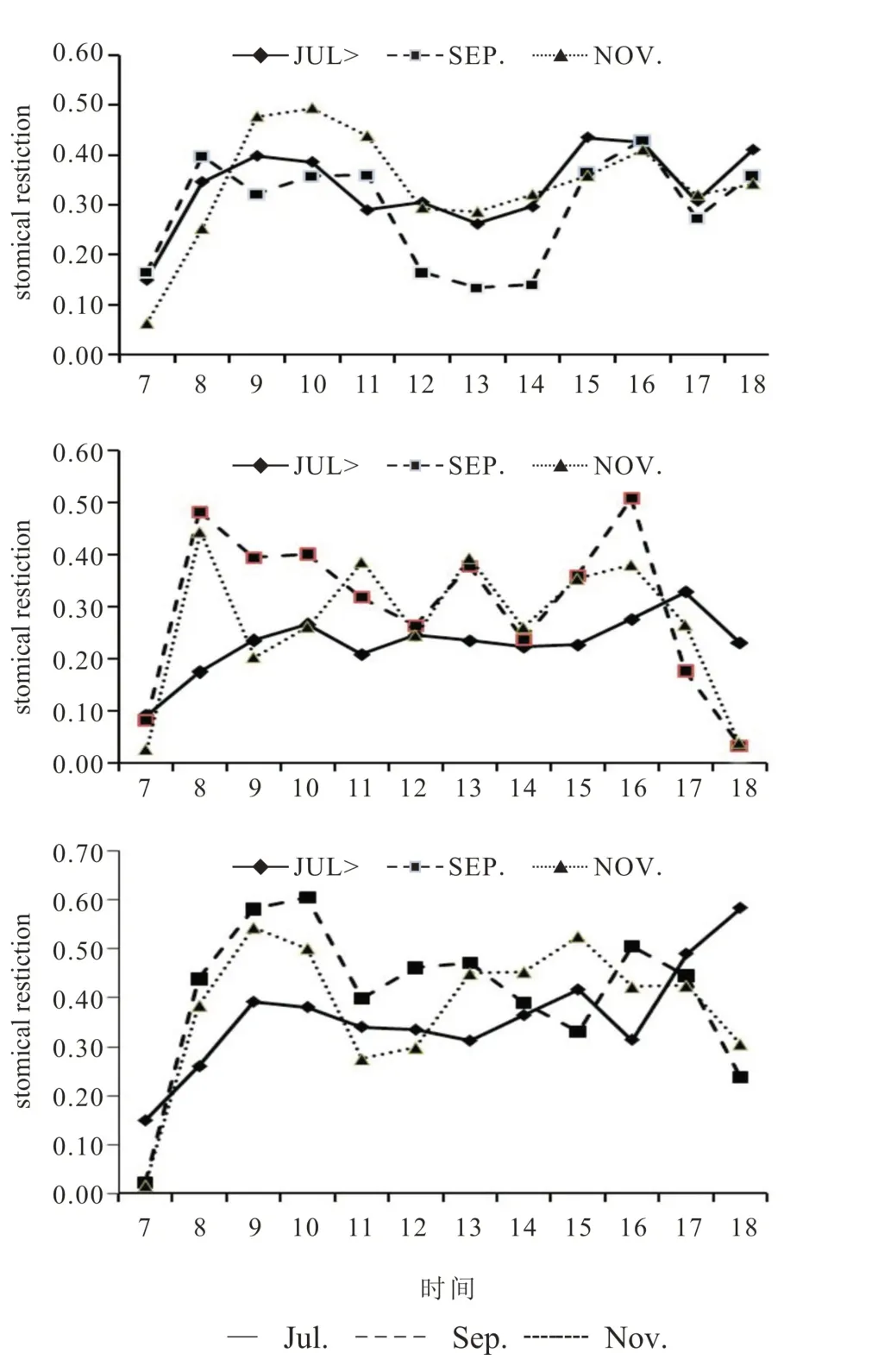
图4 气孔限制值(上有果春梢,中无果春梢下,夏梢)Fig.4 Stomatal limitation(upper psfw,,middle pspw,down psu)
2.2 光合作用的光响应曲线
叶片光补偿点10~93 μmol/(m2s),饱和光强1 372~1 776 μmol(/m2s)(表1),说明“泉口冰雪柚”利用弱光和强光能力都较强,表观量子效率为0.010 5~0.053 0 molCO2/mol photons;不同类型叶片相同月份和同类型叶片不同月份间均没有表现出明显的变化规律。

表1 饱和光强、光补偿点和表观量子效率Table 1 saturation(LSI)intensity and light compensation(LCI)and apparent quantum efficiency (AQY)
2.3 光合作用的CO2响应曲线
Pn在CO2浓度50~100 μmol/mol由负变正(表2),在0~250 μmol/mol 浓度范围内缓慢上升;在250~900 μmol/mol 范围内,随CO2浓度提高Pn剧增,在900 μmol/mol开始,即使增加CO2浓度,Pn增幅很小或不增加,甚至下降,其整个进程近似双曲线。CO2补偿点为42~92 μmol/mol,饱和点为778~1 136 μmol/mol,羧化效率0.014 6~0.048 0 mol/(m2s)。

表2 CO2的补偿点、饱和CO2浓度、羧化效率和曲线响应方程Table 2 CO2 compensation(CCC)、and saturation concentration(CSC)、carboxying efficiency(CE)and fitting response equation between Pn and air CO2 concentration
3 讨论
“泉口冰雪柚”外围枝梢叶片晴天条件下与柑桔其他种类以及其他果树的日进程[10-14]相似,呈双峰曲线,有明显“午休”现象。发生“午休”现象的原因主要是强光、高温、低湿和土壤干旱等因素,Farquhar和Sharkey把限制光合速率的因素分为气孔因素(主要是气孔导度下降导致CO2供应不足)和非气孔因素(主要是光合能力下降)两大类。本试验证明“泉口冰雪柚”的“午休”是由两种因素共同作用的结果,同类型叶片7 月“午休”弱于9 月和11 月,7 月的光强和叶片温度比9 月和11 月高,出现“午休”时气孔导度下降(高出9 月和11 月),胞间CO2浓度上升(幅度小于9月和11月)或基本不上升,这说明7 月“午休”的主要原因是气孔导度下降,气孔因素占主要地位;9月和11月平均湿度也只有35%,最小湿度只有16%,9 月和11 月饱和光强下的Pn值比7月Pn值要小得多,气孔导度也低于7月,虽然出现“午休”时气孔导度也下降,但胞间CO2浓度上升幅度大于7 月,这说明9 月和11 月“午休”原因主要是非气孔因素,但也有气孔因素的作用。生产实践中如果出现干旱,特别是在高光强和高温条件下,要注意降温、遮荫、增湿,创造适宜的生态小环境以打破或减弱“午休”的程度,以提高光能利用率和光合作用效率。高温、强光、低湿条件下气孔如何调节以及光合功能的下降进程和机理还有待进一步探求。
有果春梢叶片采果后有CO2补偿点和饱和光强升高、饱和CO2浓度呈下降趋势,这是果实对光合作用的影响,还是季节、天气的原因,或者是叶龄的关系,还有待进一步研究。
植物光合作用受各种生理生态因子共同影响,包括植株种类、品种、年龄、叶如龄、叶位,叶绿素含量与比叶重,酶系统,光的质量,CO2浓度,空气和土壤湿度、温度,土壤营养元素含量,外源激素及其他外源物等因素。而且这些因素(特别是外部因素)自身的多变以及内、外因素间的相互作用构成了光合作用影响因素的随机多变性、复杂性和不稳定性,这导致了不同叶片类型在不同季节对光合作用起支配作用的因子也不相同,这还有待进一步研究。
泉口冰雪柚不同类型叶片如春梢叶片、夏梢叶片净光合速率存在有差异,且同种叶片在不同季节光合作用影响因子也不完全相同,如何将叶片光合作用研究与生产实践如叶片管理、花果管理相结合有待进一步深入。
