南京椴天然居群的表型性状多样性及变异分析
2021-07-01严灵君岳远灏汤诗杰王欢利
严灵君, 黄 犀, 岳远灏, 汤诗杰, 王欢利
〔江苏省中国科学院植物研究所(南京中山植物园) 江苏省植物资源研究与利用重点实验室, 江苏 南京210014〕
南京椴(TiliamiquelianaMaxim.)隶属于椴树科(Tiliaceae)椴树属(TiliaLinn.),为落叶大乔木,株高可达20 m[1-2]。南京椴的主干挺拔、枝繁叶茂、花香馥郁,既是优良的行道和材用树种,也是良好的编织和造纸材料及蜜源植物;其花、叶中含有类黄酮、内酯类和芳香油等成分,具有治疗呼吸和消化疾病的药用功效[3-5]。南京椴天然居群主要分布于江苏、浙江、安徽和江西等地,相对集中或散生于落叶或常绿阔叶林中[3],是江苏省重要的乡土树种。研究结果[6-8]表明:南京椴天然居群处于生长衰退期,自然更新缺乏、生境破坏严重,个体数量不断减少,加之现有天然居群的遗传背景和多样性水平不清晰,南京椴种质资源的保护及遗传改良受到制约,因此,对南京椴种质资源进行收集和保护对其可持续应用具有重要的意义[9]。
植物的表型性状由遗传因子决定,但同时具有环境可塑性,反映了植物对环境因子的适应程度。经过长期的自然选择,植物表型性状发生不可逆的改变,并能稳定地遗传给后代,因此,对表型性状变异的研究是探究植物遗传多样性的重要途径之一。目前,对南京椴的相关研究主要集中在群落结构[6-8]、生态学[10]、个体生长规律[11]以及繁殖技术[12-15]等方面,但关于其种群表型性状多样性和遗传关系尚缺乏全面且深入的探究。
鉴于此,作者以南京椴的5个天然居群为研究对象,对叶片、种子和果序的11个表型性状进行测定,并在此基础上采用方差分析、相关性分析、主成分分析和聚类分析等多种分析方法,揭示南京椴天然居群的表型性状多样性特征和变异规律,同时在一定程度上解析南京椴的遗传多样性水平和变异状况,为南京椴种质资源的保护、评价及遗传改良奠定基础。
1 材料和方法
1.1 材料
在江苏省和安徽省南京椴的自然分布区选择5个天然居群供试。供试居群分别位于江苏省南京市牛首山、江苏省镇江市宝华山、安徽省滁州市琅琊山、安徽省安庆市鹞落坪和安徽省宿州市皇藏峪。各样地的基本概况见表1。
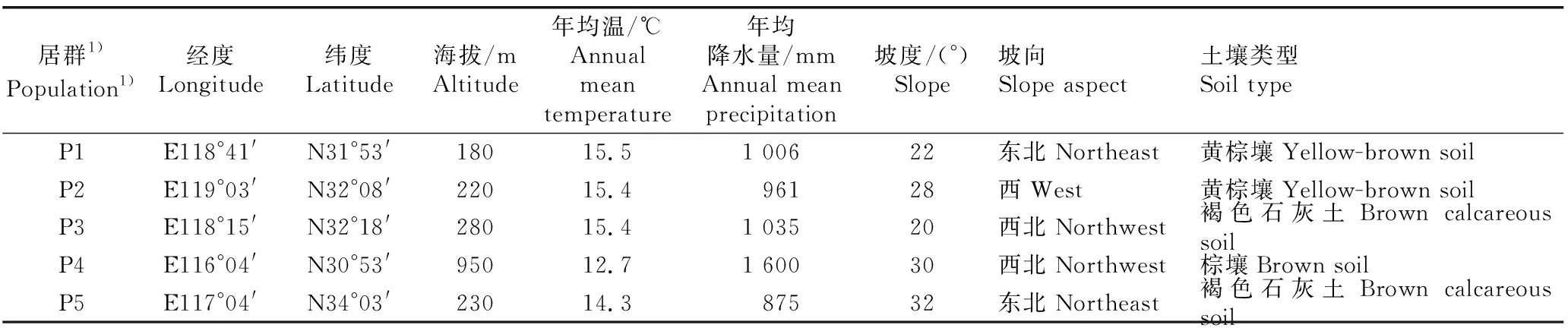
表1 供试南京椴5个天然居群的生境概况
在每个居群中选取10株生长正常、无严重缺陷、无病虫害的南京椴个体作为样株,样株树龄15~30 a、树高10 m以上、胸径18 cm以上,样株间距30 m以上。于2018年8月至10月采集样株树冠中上部南向叶片和结果小枝,用于各项表型性状的测定。
1.2 方法
从南京椴的叶片、种子和果序的形态指标中选择11个表型性状,并参照童跃伟等[16]的方法,使用游标卡尺(精度0.01 mm)对各表型性状进行精确测量。其中,叶片表型性状包括叶柄长(叶柄基部至叶片基部的长度)、叶片长(叶片基部至顶端的长度)和叶片宽(叶片最宽处的宽度);种子表型性状包括种柄长(种柄基部至种子基部的长度)、种子横径(种子横轴长度)和种子纵径(种子纵轴长度);果序表型性状包括苞片柄长(苞片柄基部至苞片基部的长度)、苞片长(苞片基部至顶端的长度)、苞片宽(苞片最宽处的宽度)、苞果合生长度(苞片上果序着生点至苞片柄顶部的长度)和果序柄长(果序柄基部至果序基部的长度)。
每个居群10株样株,采集各样株树冠中上部南向枝条5支,在枝条中上部随机采集完整的叶片、种子、苞片及果序样本各30个,每个样本各表型性状分别测量3次,结果取平均值。
1.3 数据处理和统计分析
采用EXCEL 2010软件对实验数据进行整理;并采用R3.3.3软件进行数据分析。采用箱式图进行离群值的检测,对分组数据进行正态分布和方差齐性检测。
分别对各表型性状的均值和标准差进行计算和单因素方差分析,并采用Fisher最小显著性差异法(LSD)对5个天然居群11个表型性状均值进行多重比较。
分别计算各表型性状的均值和标准差,采用变异系数(CV)分析各表型性状的离散程度[17];采用Shannon-Wiener多样性指数(H′)分析5个天然居群表型性状的多样性程度[18];参考苏泽春等[19]的方法对各表型性状进行巢式方差分析,并计算表型分化系数;采用Pearson相关系数分析各表型性状的相关性,同时采用corr.test函数对相关系数进行显著性检验;采用rda函数进行表型性状的主成分分析;将数据进行Z-score标准化处理,并采用Ward法对5个天然居群进行聚类分析。
2 结果和分析
2.1 南京椴天然居群表型性状的差异性和多样性分析
南京椴5个天然居群的叶片、种子和果序表型性状的差异、变异系数和Shannon-Wiener多样性指数的比较见表2,各表型性状的方差分析结果见表3,各表型性状的方差分量和表型分化系数见表4。
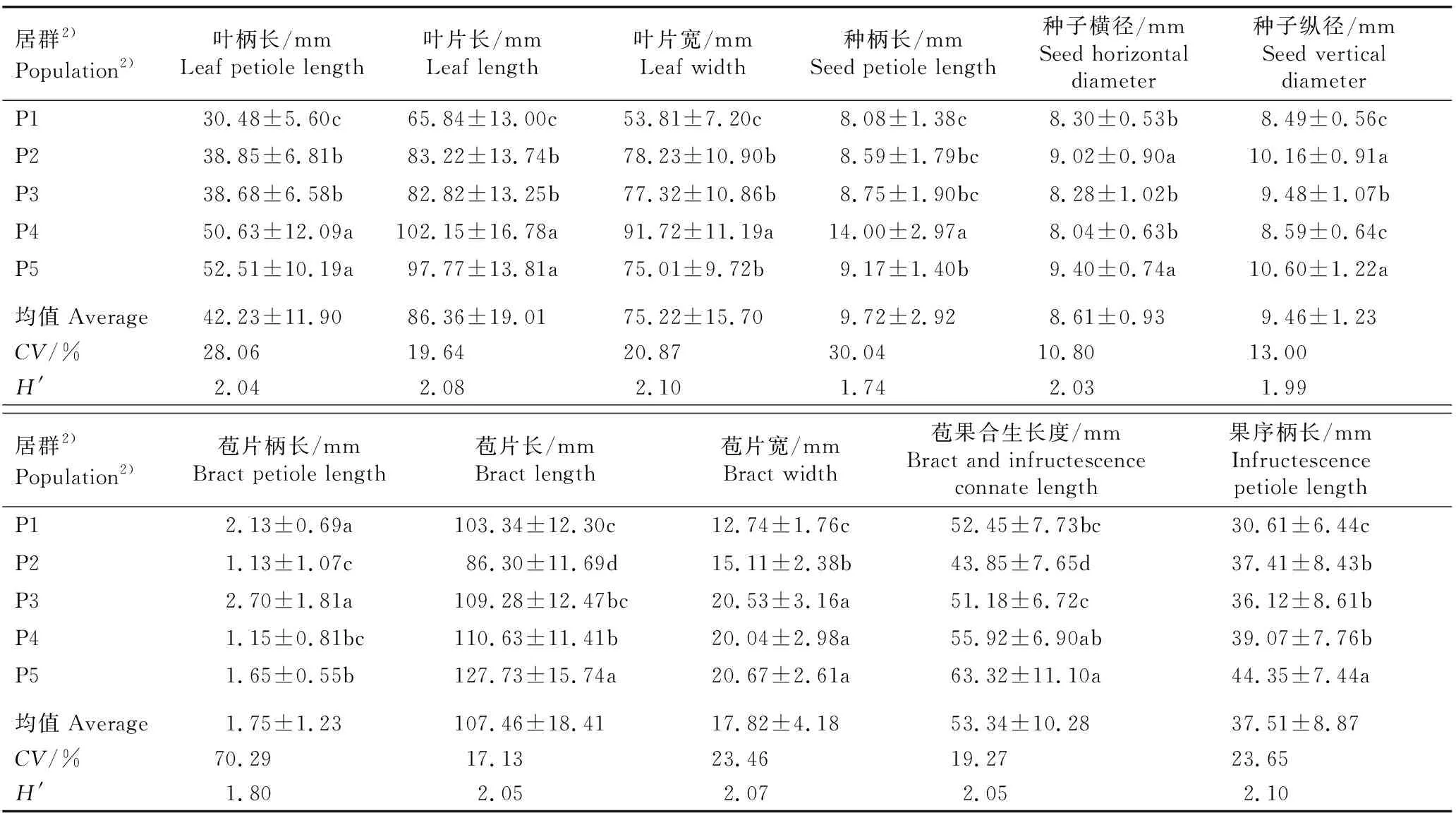
表2 南京椴5个天然居群叶片、种子和果序表型性状的差异、变异系数和Shannon-Wiener多样性指数的比较1)
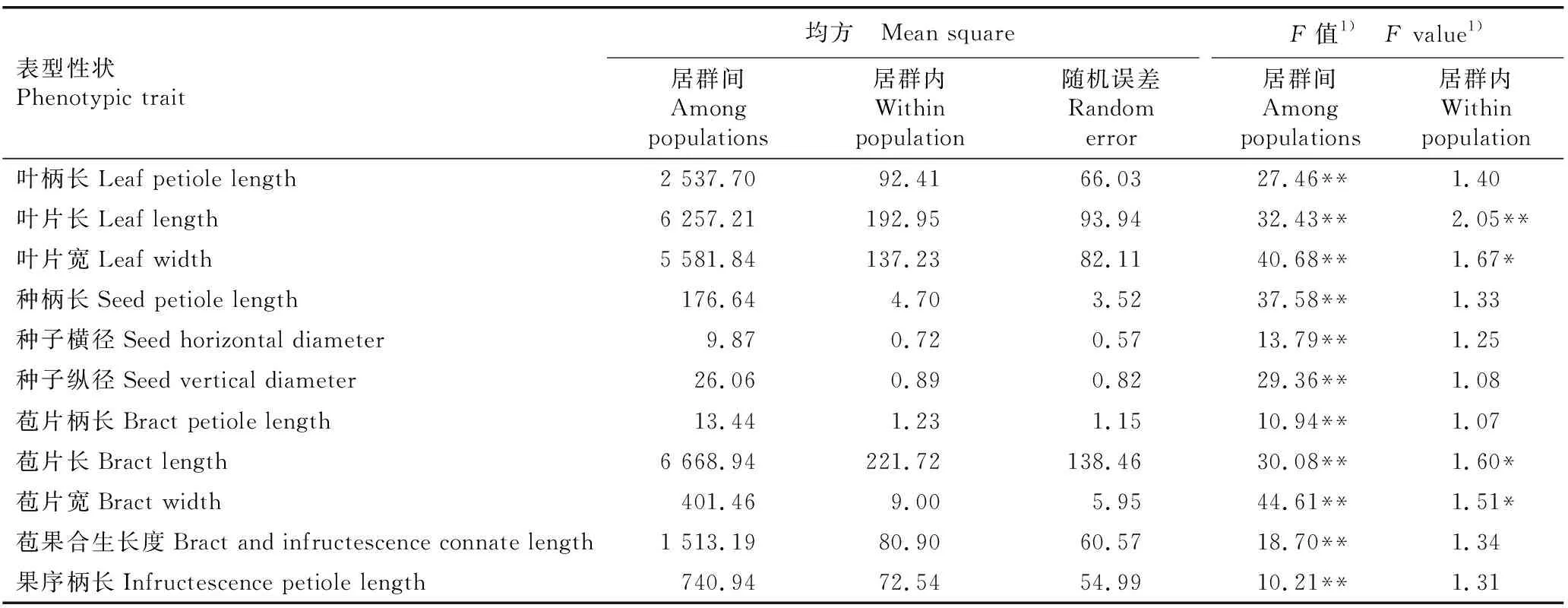
表3 南京椴居群间和居群内叶片、种子和果序表型性状的方差分析结果
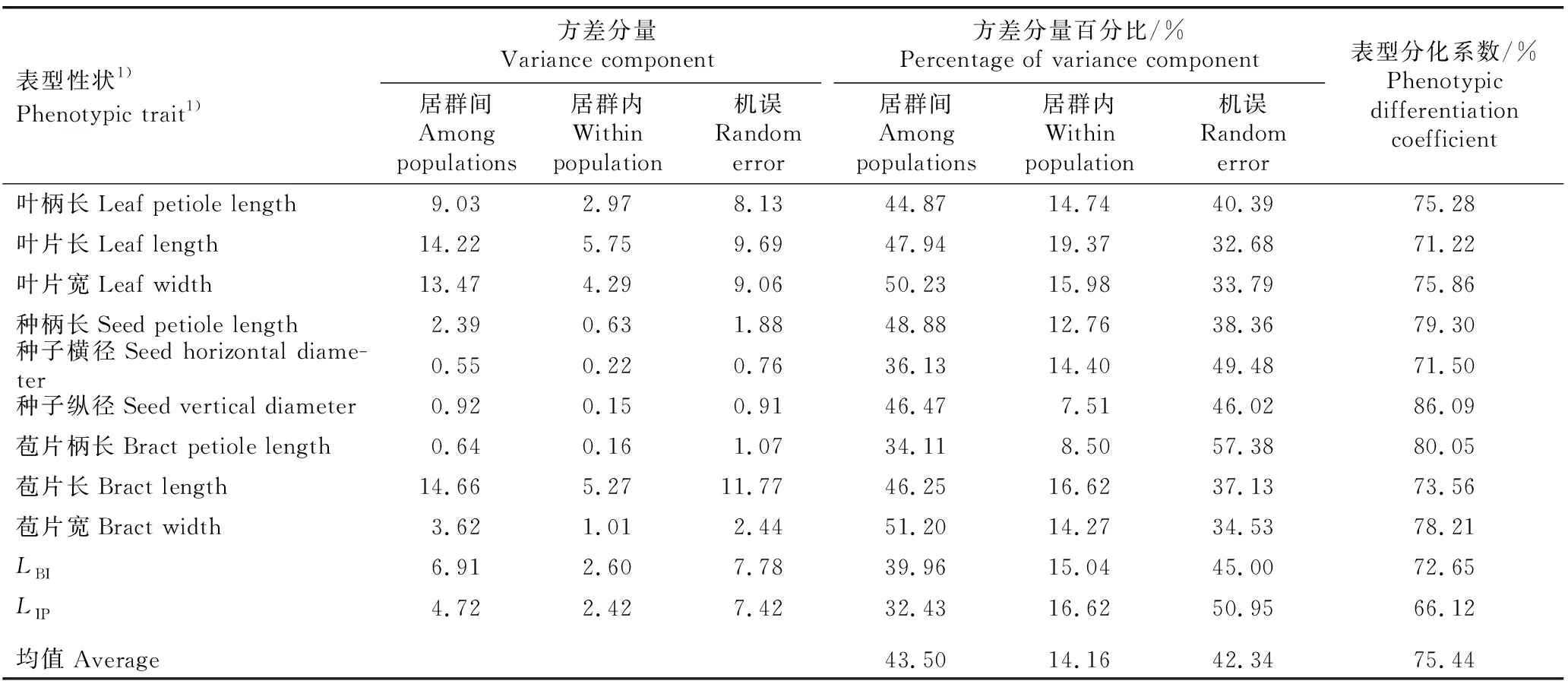
表4 南京椴天然居群叶片、种子和果序表型性状的方差分量和表型分化系数
2.1.1 表型性状的差异 由表2可见:南京椴的叶柄长、叶片长、叶片宽、种柄长、种子横径、种子纵径、苞片柄长、苞片长、苞片宽、苞果合生长度和果序柄长的均值依次为42.23、86.36、75.22、9.72、8.61、9.46、1.75、107.46、17.82、53.34和37.51 mm,不同居群间各表型性状均存在不同程度的差异。
在3个叶片表型性状中,安徽省宿州市皇藏峪(P5)居群的叶柄长(52.51 mm)最大,显著(P<0.05)大于安徽省滁州市琅琊山(P3)、江苏省镇江市宝华山(P2)和江苏省南京市牛首山(P1)3个居群;安徽省安庆市鹞落坪(P4)居群的叶片长(102.15 mm)和叶片宽(91.72 mm)均最大,且总体与其他4个居群差异显著;P1居群的叶柄长(30.48 mm)、叶片长(65.84 mm)和叶片宽(53.81 mm)均最小,且与其他4个居群差异显著。
在3个种子表型性状中,P4居群的种柄长(14.00 mm)最大,且与其他4个居群差异显著;P5居群的种子横径(9.40 mm)和种子纵径(10.60 mm)均最大,与P2居群无显著差异,但二者的种子横径和种子纵径均显著大于其他3个居群。
在5个果序表型性状中,P3居群的苞片柄长(2.70 mm)最大,与P1居群无显著差异,但二者的苞片柄长均显著大于其他3个居群;P5居群的苞片长(127.73 mm)、苞果合生长度(63.32 mm)和果序柄长(44.35 mm)均最大,且总体上与其他4个居群差异显著;P5、P4和P3居群的苞片宽分别为20.67、20.04和20.53 mm,三者无显著差异,但均显著大于P2和P1居群。
方差分析结果(表3)显示:南京椴的11个表型性状的差异在居群间均达极显著(P<0.01)水平;但在居群内仅叶片长这一表型性状的差异达极显著水平,叶片宽、苞片长和苞片宽3个表型性状的差异达显著水平,其他7个表型性状在居群内均无显著差异。
2.1.2 表型性状的变异性 由11个表型性状的变异系数(表2)可见:南京椴天然居群11个表型性状的变异系数(CV)存在较大差异,其中,种子横径的CV值最小,仅为10.80%;苞片柄长的CV值最大,达到70.29%,且总体上看,叶柄长、种柄长和苞片柄长的变异系数在各自对应的器官表型性状中均最大。
在3个叶片表型性状中,叶柄长的CV值(28.06%)最大,叶片长的CV值(19.64%)最小。在3个种子表型性状中,种柄长的CV值(30.04%)最大,种子横径的CV值(10.80%)最小。在5个果序表型性状中,苞片柄长的CV值(70.29%)最大,苞片长的CV值(17.13%)最小,果序柄长、苞片宽和苞果合生长度的CV值均在24%以下。
总体上看,在南京椴天然居群的11个表型性状中,仅种子横径和种子纵径的CV值小于15%,其他9个表型性状的CV值均在15%以上,说明南京椴天然居群表型性状的变异较为丰富,但种子表型性状的变异程度小于其他器官。
2.1.3 表型性状的多样性 由Shannon-Wiener多样性指数(表2)可见:南京椴5个天然居群11个表型性状的Shannon-Wiener多样性指数(H′)存在一定差异,其中,种柄长的H′值(1.74)最小,叶片宽和果序柄长的H′值(2.10)最大,且总体上叶片性状的H′较大且变化幅度较小。
在3个叶片表型性状中,叶片宽的H′值(2.10)最大,叶柄长的H′值(2.04)最小。在3个种子表型性状中,种子横径的H′值(2.03)最大,种柄长的H′值(1.74)最小。在5个果序表型性状中,果序柄长的H′值(2.10)最大,苞片宽、苞片长和苞果合生长度的H′值均在2.05及以上,而苞片柄长的H′值(1.80)最低。说明供试南京椴天然居群的种子表型性状的多样性较低,叶片和果序的表型性状多样性较高。
2.1.4 表型性状的变异来源 由表4可见:在南京椴天然居群11个表型性状的居群间方差分量百分比的均值(43.50%)明显大于居群内方差分量百分比的均值(14.16%)。各表型性状的居群间表型分化系数为66.12%~86.09%,均值为75.44%;其中,仅果序柄长的表型分化系数为66.12%,其他表型性状的表型分化系数均在70%以上。说明居群间的变异是供试南京椴表型性状变异的主要来源。
2.2 南京椴天然居群表型性状的相关性分析
南京椴天然居群的叶片、种子和果序表型性状间的相关系数见表5。
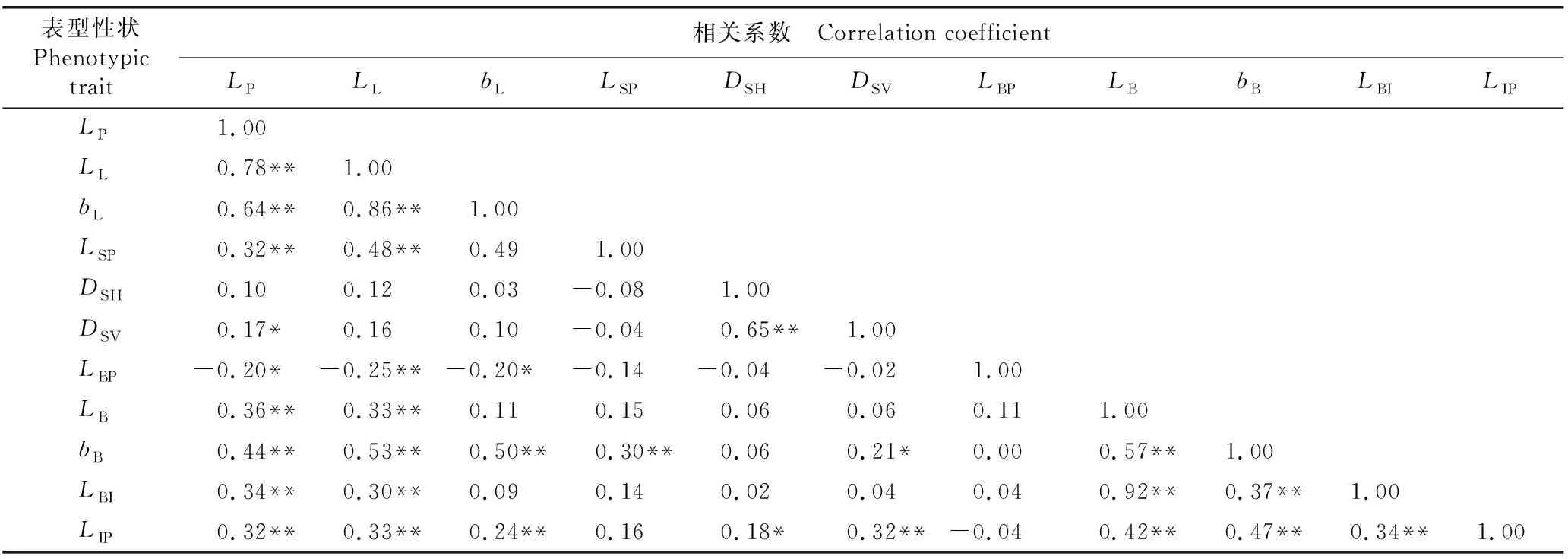
表5 南京椴天然居群的叶片、种子和果序表型性状间的相关系数1)
由表5可见:3个叶片表型性状(叶柄长、叶片长和叶片宽)间均呈极显著(P<0.01)正相关;而在3个种子表型性状中,仅种子横径与种子纵径呈极显著正相关;在5个果序表型性状中,苞片长、苞片宽、苞果合生长度和果序柄长4个表型性状间呈极显著正相关。而在供试的11个表型性状中,叶柄长仅与种子横径无显著相关性,叶片长仅与种子横径和种子纵径无显著相关性,且种子的3个表型性状与多数果序表型性状间均无显著相关性;另外,苞片柄长与3个叶片表型性状呈显著(P<0.05)或极显著负相关,与所有种子表型性状和果序表型性状均无显著相关性。
总体上看,种子表型性状与叶片和果序表型性状间大多无显著相关性,而叶片表型性状与果序表型性状间大多具有极显著或显著相关性。
2.3 南京椴天然居群表型性状的主成分分析
对南京椴天然居群11个表型性状进行主成分分析,结果见表6。结果显示:前5个主成分的累计贡献率达82.96%,表明前5个主成分可基本反映南京椴表型性状的大部分信息。
第1主成分的贡献率为36.35%,以叶柄长、叶片长、叶片宽、苞片长、苞片宽、苞果合生长度和果序柄长特征向量的绝对值较大,均在1.0以上;第2主成分的贡献率为16.58%,以叶片宽、苞片长和苞果合生长度特征向量的绝对值较大,均在1.0以上,说明第1和第2主成分主要反映南京椴叶片和苞片等器官的特征。第3主成分的贡献率为15.21%,以种子纵径和种子横径特征向量的绝对值较大,均在1.5以上,说明该主成分主要反映南京椴种子的特征。第4主成分的贡献率为8.22%,以苞片柄长特征向量的绝对值较大(大于1.5),说明该主成分主要反映南京椴苞片柄的特征。第5主成分的贡献率为6.60%,以果序柄长特征向量的绝对值较大(大于1.0),说明该主成分主要反映南京椴果序柄的特征。
2.4 南京椴天然居群的聚类分析
基于叶片、种子和果序11个表型性状对南京椴5个天然居群进行聚类分析,结果见图1。
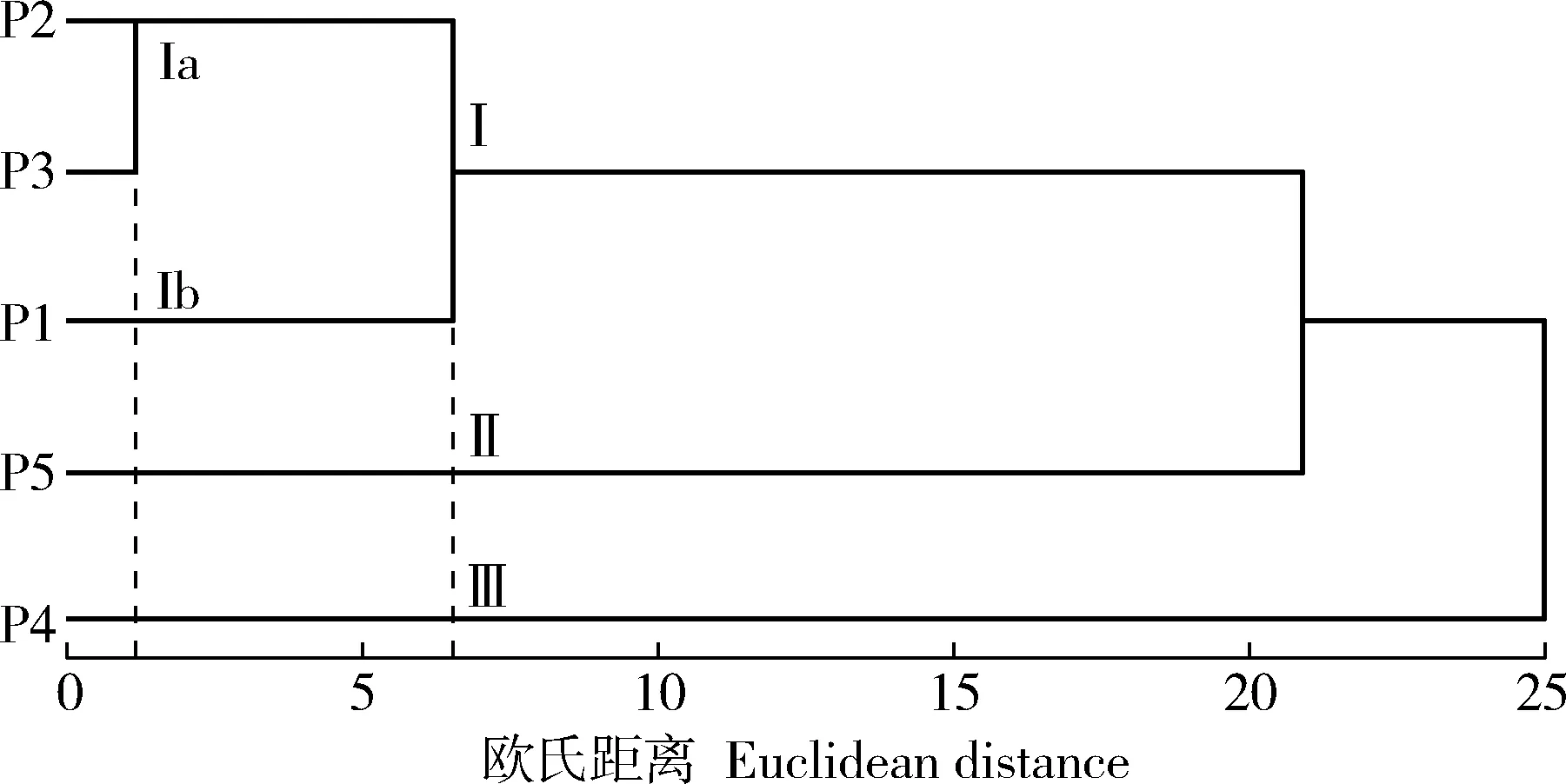
P1: 江苏省南京市牛首山Niushou Mountain of Nanjing City in Jiangsu Province; P2: 江苏省镇江市宝华山Baohua Mountain of Zhenjiang City in Jiangsu Province; P3: 安徽省滁州市琅琊山Langya Mountain of Chuzhou City in Anhui Province; P4: 安徽省安庆市鹞落坪Yaoluoping of Anqing City in Anhui Province; P5: 安徽省宿州市皇藏峪Huangcangyu of Suzhou City in Anhui Province.
在欧氏距离6.0处,供试的5个天然居群可分为3组,其中,江苏省镇江市宝华山(P2)、安徽省滁州市琅琊山(P3)和江苏省南京市牛首山(P1)3个居群聚在一起,为组Ⅰ;安徽省宿州市皇藏峪(P5)和安徽省安庆市鹞落坪(P4)居群各自单独成组,分别为组Ⅱ和组Ⅲ。在欧氏距离1.0处,组Ⅰ又可分为2个亚组,其中,P2和P3居群聚为亚组Ⅰa,P1居群单独成亚组Ⅰb。从表型性状看,P2和P3居群的叶柄长、叶片长、叶片宽、种柄长、种子横径、种子纵径以及果序柄长的测定值均较为相近;P1居群的叶柄长、叶片长、叶片宽和种柄长均明显小于P2和P3居群;P4和P5居群的叶片表型性状(叶柄长、叶片长和叶片宽)以及种柄长均明显大于其他居群。
供试南京椴5个居群的聚类分析结果与各居群的地理距离有一定关系。江苏省南京市牛首山、江苏省镇江市宝华山和安徽省滁州市琅琊山3个居群的地理距离较近,在聚类图上聚为一组;而安徽省安庆市鹞落坪和安徽省宿州市皇藏峪2个居群与上述3个居群的地理距离较远,在聚类图中各自成组。
3 讨论和结论
植物的表型性状既具有变异性又具有稳定性,受其自身的遗传组成和外部环境两方面的影响,可在一定程度上直观地反映物种的遗传多样性[20-21]。南京椴5个天然居群的叶片、种子和果序的表型性状在居群内和居群间均存在一定的差异,其中,居群间11个表型性状的差异达到极显著水平,而在居群内仅叶片长、叶片宽、苞片长和苞片宽存在极显著或显著差异,且11个表型性状的居群间表型分化系数均值为75.44%,表明南京椴天然居群的表型性状变异较大,且变异主要来自居群间;而汤诗杰等[22-23]基于ISSR和RAPD遗传标记对南京椴天然居群的遗传变异状况进行了分析,认为居群内变异是南京椴遗传分化的主要来源。南京椴的表型性状分析与遗传标记分析的结果存在差异,这一现象说明表型性状不仅由遗传因子决定,同时还存在一定的可塑性,尤其是营养器官的形态,受不同区域内土壤、水分和植被类型等因子影响,是植物适应环境因子的一种表现。
南京椴11个表型性状的变异系数为10.80%~70.29%,表明南京椴表型性状的变异丰富;且除种子横径和种子纵径外,其他表型性状的变异系数均在15%以上,表明与营养器官相比,种子等繁殖器官受环境变化的影响较小,性状的稳定性相对较强。11个表型性状的Shannon-Wiener多样性指数为1.74~2.10,表明南京椴的表型性状具有丰富的多样性;其中,果柄长和叶片宽的Shannon-Wiener多样性指数最高,种柄长的Shannon-Wiener多样性指数最低,与各自变异系数的分析结果存在一定差异。形成这种差异的原因可能与这2类数据蕴涵的意义不同有关。变异系数反映了表型性状数据的离散程度;而Shannon-Wiener多样性指数则是对表型性状丰富度和均匀度的综合评价,表型性状越丰富且分布越均匀,多样性指数越高[24]。
相关性分析结果表明:南京椴的叶片、种子和果序3类器官的各表型性状间存在不同程度的相关性,叶片和果序表型性状与种子横径和纵径总体无显著相关性,但叶片表型性状与果序表型性状大多呈极显著或显著相关性,说明同类器官的表型性状存在一定的相关性,但不同类器官间的表型性状关联度不高,特别是种子表型性状与其他2类器官多数表型性状均无显著相关性,而叶片表型性状与果序表型性状(特别是苞片表型性状)大多存在一定的相关性,这与苞片(变态叶)与叶片的器官属性一致有关。
对南京椴表型性状的主成分分析结果表明:前5个主成分的累计贡献值达82.96%,其中第1、第2、第4和第5主成分主要反映南京椴叶片和果序的特征,第3主成分主要反映其种子的特征。前5个主成分中绝对值较大的表型性状均为反映南京椴表型性状差异的主要因子,也是南京椴多样性保护和选育研究中的主要观测性状。聚类分析结果表明:在欧氏距离6.0处,可将供试的5个南京椴天然居群分为3组,其中,地理分布较近的P1、P2和P3居群聚为一组,这3个居群分别分布于江苏省南京市牛首山、江苏省镇江市宝华山和安徽省滁州市琅琊山,而地理距离相对较远的P4和P5居群则各自独立成组,这2个居群分别分布于安徽省安庆市鹞落坪和安徽省宿州市皇藏峪,表明基于表型性状的聚类结果与各居群的地理分布基本一致。而汤诗杰等[22-23]基于分子标记对南京椴天然居群进行的聚类分析结果并未表现出居群遗传因子和地理分布的一致性。植物的表型特征不仅由其基因型决定,也受环境压力的影响[25],这是导致南京椴居群遗传分化与表型分化规律不一致的原因之一。
综上所述,南京椴天然居群的表型性状变异丰富,不同居群的种子及叶片等器官的形态存在丰富的多样性;居群间的表型分化是其变异的主要来源。植物的表型性状存在一定的环境可塑性,是其适应环境因子的一种表现。植物的天然居群保持较高的遗传多样性水平有利于其居群的发展[26],因此,在南京椴种质资源的保护过程中,可选取表型差异较大的个体进行重点保护,通过对表型性状的聚类分组阐明不同居群表型性状的差异,并依据育种目标筛选出适宜的育种材料。
本研究选取的5个天然居群主要分布于江苏省和安徽省,地理位置相近且生境条件相似,获得的研究数据不能充分解析环境因子对表型性状的影响,因而,后续可进一步根据南京椴的分布区域扩大取样范围,通过分析不同分布区的气候和土壤因子与表型性状的相关性,解析其表型性状的遗传稳定性和环境可塑性。此外,本研究仅对南京椴的叶片、种子和果序等器官的表型性状进行了1年定点测量,存在表型性状较少、观察和测量连续性和重复性不足等问题,因而,后续可以通过连年取样或建立同质园实验,扩大样本数量并增加不同组织和器官表型性状的数量,以期更全面地反映南京椴天然居群的表型特征差异。在今后的研究中,还可以对南京椴分布区域的环境因子进行动态监测,结合不同居群的表型特征建立相关矩阵,进一步阐述表型特征对环境因子的响应;同时,采用分子生物学技术深入系统地揭示南京椴天然居群的遗传变异规律,以期推动南京椴天然居群的遗传多样性研究及其野生资源的保护、评价和利用。
