基于花器官形态特征的广义李属植物的数量分类
2021-07-01赵旭明吴保欢王永刚崔大方
赵旭明, 吴保欢, 王永刚, 李 薇, 崔大方,①
(1. 华南农业大学: a. 林学与风景园林学院, b. 华南农业博物馆, 广东 广州 510642; 2. 中山市小榄中学, 广东 中山 528000; 3. 新疆农业科学院农作物品种资源研究所, 新疆 乌鲁木齐 830091)
广义李属(PrunusLinn.sensulato)植物隶属于蔷薇科(Rosaceae)李亚科(Subfam. Prunoideae Focke),为核果类落叶或常绿乔、灌木,全球有200~250种,广泛分布于北半球[1]。广义李属植物生境多样,从气候干燥的温带地区到较湿润的亚热带、热带地区,从寒冷的高山地带到气候温和的低海拔区域,均有分布[2]。1700年,Tournefort[3]基于果实形态将这类核果类群分为6个属级单位:PrunusLinn.sensustricto、ArmeniacaMill.、CerasusMill.、AmygdalusLinn.、PersicaMill.和LaurocerasusTourn ex Duh.。1865年,Bentham等[4]首先提出广义李属的概念,将蔷薇科所有核果类植物合并为广义李属,并在属下划分7个组。1893年,Koehne[5]将这7个组划分为7个亚属。1940年,Rehder[6]基于花序和内果皮特征将广义李属划分为除臭樱属(MaddeniaJ. D. Hook. et Thoms.)和臀果木属(PygeumGaertn.)以外的李亚属(Subg.PrunusLinn.)、桃亚属〔Subg.Amygdalus(Linn.) Focke〕、樱亚属〔Subg.Cerasus(Mill.) A. Gray〕、稠李亚属〔Subg.Padus(Mill.) Focke〕和桂樱亚属〔Subg.Laurocerasus(Tourn. ex Duh.) Rebd.〕,并在樱亚属下设立矮樱组(Sect.MicrocerasusKoehne)。300多年来,李亚科植物的分类一直是“分而复合,合而再分”。黄文鑫等[7]认为,广义李属的分类问题主要集中在李属的范围不确定、属下类群分类处理迥异以及部分种的分类处理仍然存在争议3个方面。
随着分子系统学的深入研究[8-15],学者们发现广义李属在系统发育树上均非单系群,认为广义李属所包含的范围和类群有:落叶单花类群,包括桃〔Prunuspersica(Linn.) Batsch〕、李(PrunussalicinaLindl.)和杏(PrunusarmeniacaLinn.)等;落叶伞形花序类群,包括樱亚属但不包括矮樱类〔Subg.Cerasus(Mill.) A. Gray Grex I.MicrocerasusC. K. Schneid.〕;总状花序类群,包括稠李(PrunuspadusLinn.)、桂樱(PrunuslaurocerasusLinn.)、臭樱(MaddeniahypoleucaKoehne)和臀果木(PygeumtopengiiMerr.)等。鉴于此,本研究基于广义李属及相近类群共47种(含变型)植物的花器官形态特征进行观察和聚类分析,通过比较研究寻求有分类意义的花器官形态特征,旨在进一步为广义李属植物的分类和系统演化提供依据。
1 材料和方法
1.1 材料
在野外观察和标本采集的基础上,以蔷薇科李亚科广义李属及相近类群共47种植物(表1)为研究材料,凭证标本保存于华南农业大学标本馆(CANT)。

表1 广义李属及相近类群植物凭证标本信息
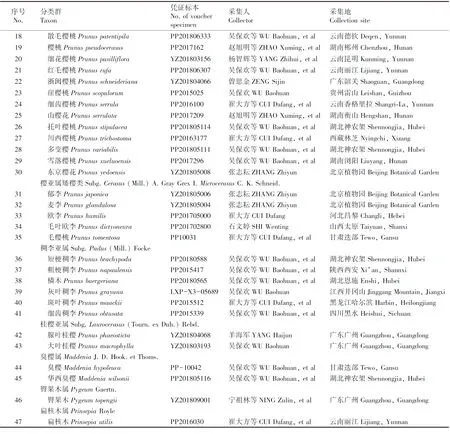
续表1 Table 1 (Continued)
1.2 方法
参照《中国植物志》[16]核对广义李属及相近类群植物的花器官形态描述,最终确定64个形态特征进行观察和测量,其中数量性状24个,定性性状40个(表2)。数量性状采用测量尺(精度0.5 mm)或游标卡尺(精度0.02 mm)测量,每个性状测量5次,计算平均值。定性性状采用肉眼和放大镜观察。以性状表现在观察统计中超过60%为准。测量和信息采集选择盛花期或有成熟花朵的标本,花器官保存较好,特征性状完整。

表2 广义李属及相近类群植物花器官形态特征及定性性状编码

续表2 Table 2 (Continued)
1.3 数据计算及分析
基于64个花器官形态特征数据,采用NTSYSpc version 2.10e软件对广义李属及相近类群植物进行UPGMA聚类分析和主成分分析。其中,所得数据矩阵用Similarity模块的Interval data命令计算,得到欧氏距离矩阵,最后用Clustering模块的Shan命令按UPGMA方法对所得欧氏距离矩阵进行聚类分析;主成分分析中用Similarity模块的Interaval data命令对标准化的数据矩阵计算相关系数,用Dcenter命令对相关系数矩阵去中心化处理,最后用Ordination模块的Eigen命令计算特征值和特征向量。
2 结果和分析
2.1 花器官形态特征及聚类分析
供试广义李属及相近类群植物的花序有总状花序、伞房状总状花序、伞形花序以及花单生或2(3)朵簇生4种类型(图1)。其中,总状花序有稠李亚属、桂樱亚属、臭樱属和臀果木属,伞形或伞房状总状花序有樱亚属(除矮樱类),花单生或2(3)朵簇生有李亚属和樱亚属的矮樱类,花单生有杏亚属〔Subg.Armeniaca(Mill.) Nakai〕和桃亚属。

1: 总状花序(细齿稠李)Raceme (Prunus obtusata Koehne); 2: 伞房状总状花序(锥腺樱桃)Corymb raceme (Prunus conadenia Koehne); 3: 伞形花序(尾叶樱桃)Umbel (Prunus dielsiana C. K. Schneid.); 4: 花单生(山杏)Solitary flower 〔Prunus sibirica (Linn.) Lam.〕; 5: 花2朵簇生(郁李)2 fascicled flowers (Prunus japonica Thunb.).
从花序梗长来看,总状花序类的花序梗长大于伞形花序类和伞房状总状花序类,而花单生或2(3)朵簇生类无花序梗。总状花序类中稠李亚属除斑叶稠李(PrunusmaackiiRupr.)外,其他稠李亚属种类花序梗长基本大于100 mm;总状花序类中桂樱亚属和臭樱属的花序梗长20~35 mm,臀果木属的花序梗长一般小于10 mm。伞形或伞房状总状花序类的樱亚属(除矮樱类)中,伞房状总状花序的散毛樱桃(PrunuspatentipilaHand.-Mazz.)花序梗长达50~80 mm,而同样分布于中国西南和西北地区的伞房状总状花序的锥腺樱桃(PrunusconadeniaKoehne)花序梗长约30 mm,其他伞房状总状花序类的花序梗长约10 mm,伞形花序类的花序梗长一般小于10 mm,甚至小于5 mm。桃亚属、李亚属、杏亚属和矮樱类的花均为单生或2(3)朵簇生,无花序梗。
从花直径来看,总状花序类中稠李亚属、桂樱亚属、臭樱属和臀果木属的花直径均小于10 mm;而伞形或伞房状总状花序类中樱亚属(除矮樱类)以及花2(3)朵簇生类中李亚属和矮樱类的花直径均在10 mm以上,其中,李亚属的花直径一般在10~20 mm,而樱亚属(除矮樱类)的花直径则在10~35 mm。
稠李亚属和臭樱属的花序梗基部有大型总苞;樱亚属(除矮樱类)的总苞较小,多为卵形或倒卵形;李亚属极少出现总苞;桂樱亚属和臀果木属无总苞。
基于64个花器官形态特征对供试广义李属及相近类群植物进行聚类分析,结果(图2)显示:在欧氏距离10.40处,47种广义李属及相近类群植物分为A、B、C、D、E、F、G和H 8个分支。A分支进一步分为A1和A2 2个分支,其中,李亚属与樱亚属的矮樱类聚为A1分支;A2分支中,毛樱桃(PrunustomentosaThunb.)和榆叶梅(PrunustrilobaLindl.)先聚在一起,然后又与杏亚属和桃聚在一起,这些植物在花器官形态上都表现为花单生或2(3)朵簇生。樱亚属(除矮樱类)植物聚为B分支;2种臭樱属植物聚为C分支,明显地聚在广义李属大类中;腺叶桂樱〔Prunusphaeosticta(Hance) Maxim.〕和稠李亚属组成D和E分支。而大叶桂樱(PrunusmacrophyllaSieb. et Zucc.)、臀果木和扁核木(PrinsepiautilisRoyle)分别独立聚为F、G和H分支。
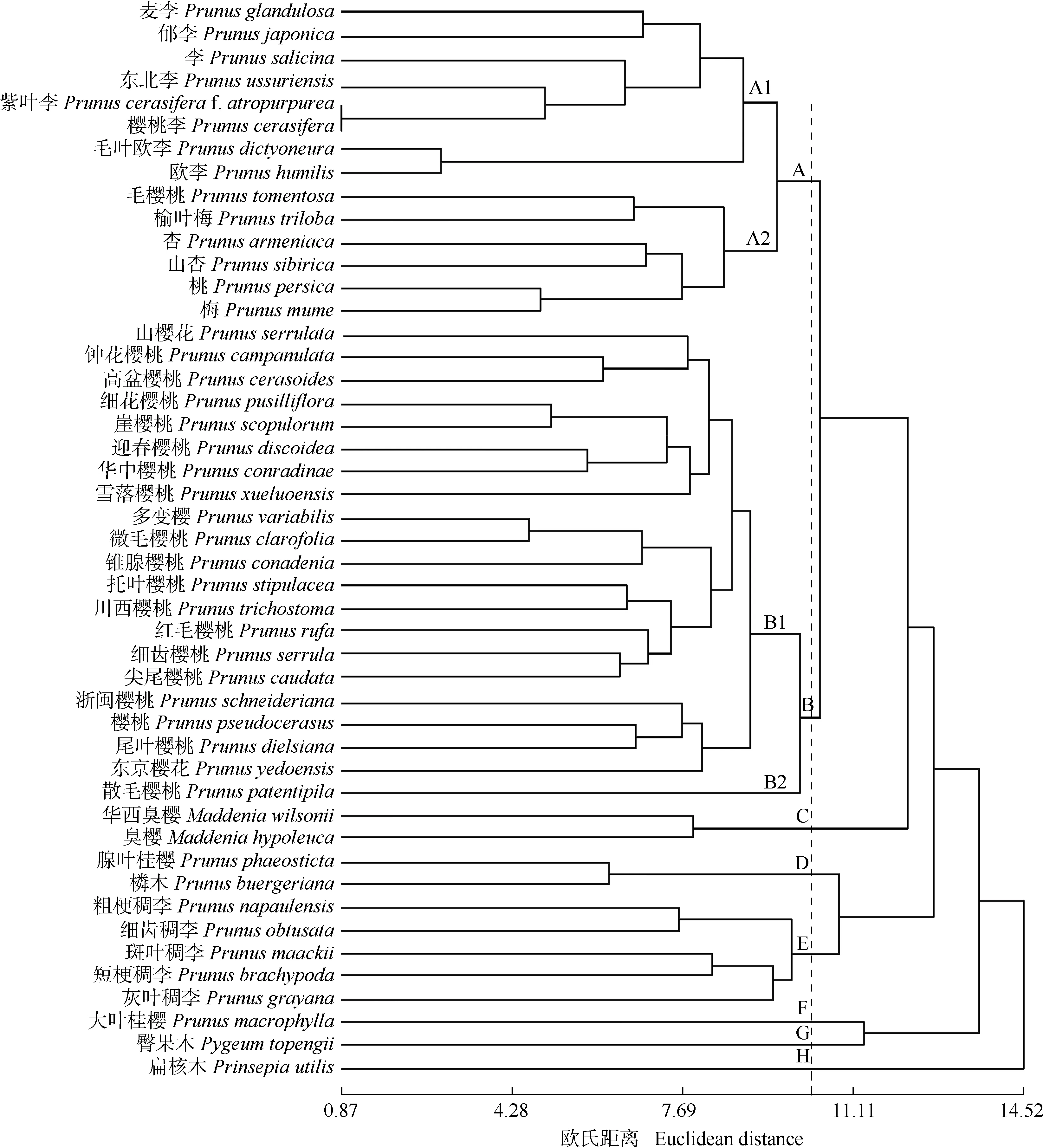
图2 基于花器官形态特征的广义李属及相近类群植物的UPGMA聚类图
2.2 主成分分析
对广义李属及相近类群植物64个花器官形态特征进行主成分分析,结果(表3)显示:前4个主成分的贡献率分别为18.764 9%、13.062 6%、8.531 7%和6.264 9%,累计贡献率仅46.624 1%;前10个主成分的累计贡献率仅超过70%,前20个主成分的累计贡献率才达到90%,累计贡献率增加非常缓慢。表明在64个花器官形态特征中,形态变异的多向性比较明显,同一形态特征可能在不同类群中都有反映,若以主成分特征对广义李属植物进行分析,至少需要10个主成分才能将核果类植物各类群区分开,这种形式会导致信息有较大的丢失,因此没有进一步的分析可行性。
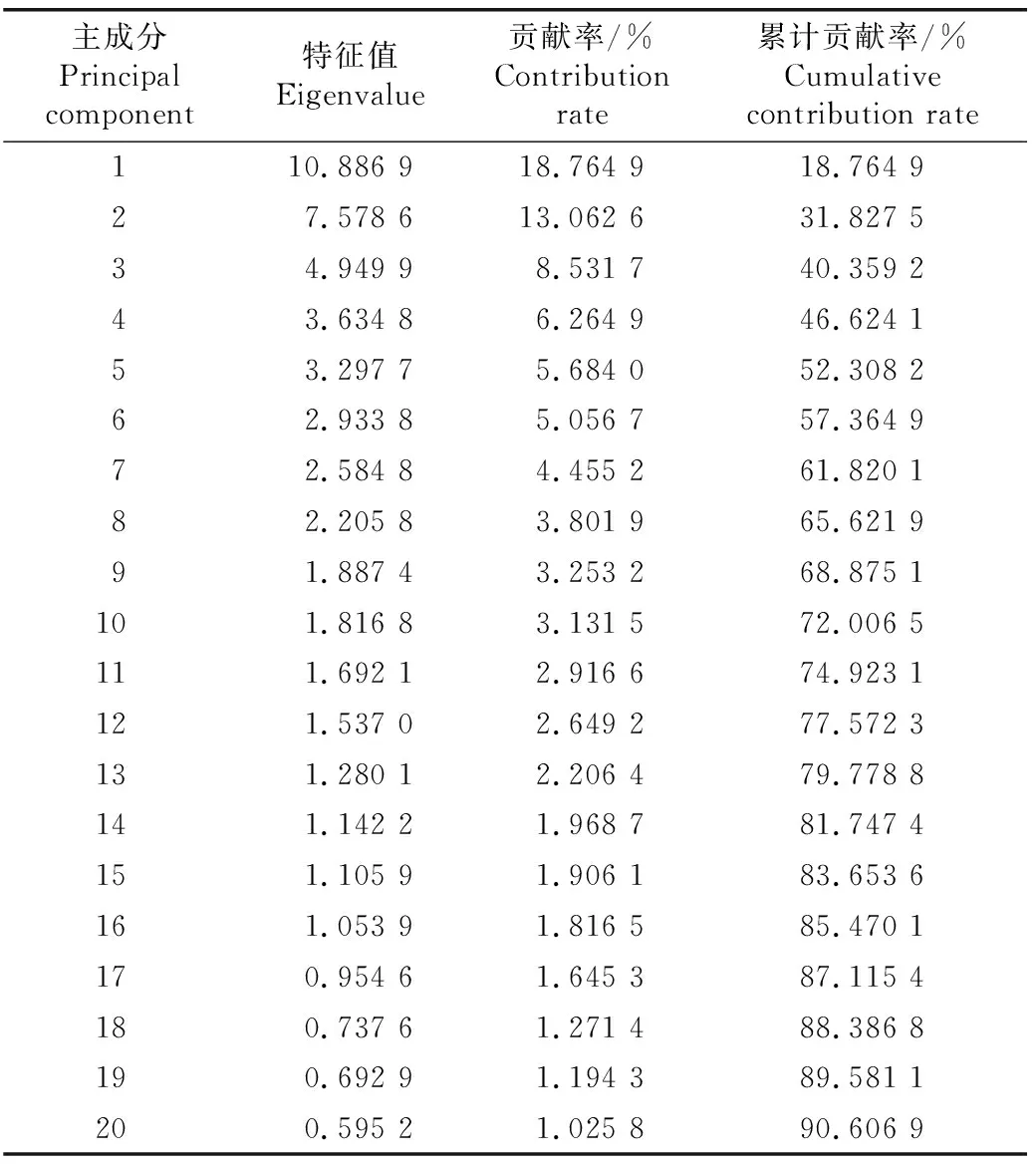
表3 广义李属及相近类群植物花器官形态特征主成分分析结果
3 讨 论
3.1 广义李属植物花器官形态的分类学意义
基于广义李属及相近类群植物64个花器官形态特征的分析结果可以看出:具有总状花序的稠李亚属、桂樱亚属、臭樱属和臀果木属能够与伞形或伞房状总状花序的樱亚属(除矮樱类)区分开,与花单生或2(3)朵簇生的桃亚属、李亚属、杏亚属和矮樱类更易区分开。另外,花序梗长、花直径以及总苞大小和形状的差异可以作为划分属内各类群的分类依据,如稠李亚属和臭樱属花序梗基部有大型总苞,花直径相对较小;樱亚属(除矮樱类)有较小的卵形或倒卵形的总苞,花直径相对较大;李亚属、桂樱亚属和臀果木属极少出现总苞或无总苞。
桃类群、李类群和杏类群常被不同的学者处理为独立的属或亚属,或将李类群和杏类群合并放入李亚属内,区分为李组(Sect.Prunus)和杏组(Sect.Armeniaca)[17-18]。聚类分析结果显示:杏亚属和桃亚属与毛樱桃聚在一起后再与李亚属及矮樱类其他成员聚为分支A,这些植物的花均为花单生或2(3)朵簇生,花瓣先端圆钝;其中李亚属、杏亚属和桃亚属花器官形态更相似,这3个亚属的花直径和花瓣较大,且子房被毛,花梗更加粗短;榆叶梅和毛樱桃形态较接近,花2(3)朵簇生,花直径约20 mm,萼片边缘具腺体,花瓣均为近圆形,二者的花柱无毛,子房密被毛。分子系统学研究[12,15]表明:桃亚属、李亚属和杏亚属关系较近,应归为同一亚属。
Koehne[19]、《中国植物志》[16]和《Flora of China》[1]都将矮樱类植物归入樱属(CerasusMill.)。聚类分析结果显示:矮樱类中花梗较长且无被毛的麦李(PrunusglandulosaThunb.)和郁李(PrunusjaponicaThunb.)聚在一起后,又与李亚属的李、东北李(PrunusussuriensisKov. et Kost.)、樱桃李(PrunuscerasiferaEhrhart)和紫叶李〔Prunuscerasiferaf.atropurpurea(Jacq.) Rehd.〕聚在一起,然后再与花梗较短的欧李(PrunushumilisBunge)和毛叶欧李(PrunusdictyoneuraDiels)聚为分支A1,并没有与樱亚属聚在一起,且与其关系较远。Shimada等[20]的杂交实验研究中,矮樱类植物与李亚属和部分桃亚属植物可以杂交,与樱亚属植物杂交则通常以失败告终,表明矮樱类与樱亚属关系较远;叶脉序[7]和分子系统学[10,14]研究结果均支持将矮樱类从樱亚属移出归入李亚属,作为李亚属下的矮樱组〔Subg.Prunussect.Microcerasus(Spach) C. K. Schneid〕。
1965年,Kalkman[21]将广义李属的范围扩大,将在热带分布的臀果木属归入桂樱亚属内。分子系统学证明,臭樱属和臀果木属嵌在广义李属中[22-23]。本研究聚类分析结果显示:臀果木和扁核木分别聚为独立的分支,这2个类群与广义李属关系较远;但臭樱和华西臭樱(MaddeniawilsoniiKoehne)聚为一分支后,又聚在广义李属大类中,其总状花序或稀有伞房花序及苞片早落、花梗短、萼筒钟状等形态学特征与桂樱亚属接近,支持将臭樱属归入广义李属中[23]。
3.2 部分种的分类处理
崖樱桃(PrunusscopulorumKoehne)、樱桃(PrunuspseudocerasusLindl.)、细花樱桃(PrunuspusillifloraCard.)和华中樱桃(PrunusconradinaeKoehne)形态相似,不易区分,但《中国植物志》[16]还是保留各种。《Flora of China》[1]将崖樱桃并入樱桃。吴保欢等[24]将细花樱桃、崖樱桃和华中樱桃合并。本研究的聚类分析结果显示:细花樱桃和崖樱桃聚在一起,且与华中樱桃的距离非常接近,在花器官形态特征上,花序均为伞形或伞房状总状花序,总苞均为椭圆形,边缘具有圆锥状腺体,萼片均为宽卵状三角形,萼筒钟状,长约为宽的2倍,花冠颜色均为白色,花柱与雄蕊的高度几一致,而樱桃因其花梗密被毛与细花樱桃、崖樱桃和华中樱桃区分开来,本文支持将崖樱桃和细花樱桃并入华中樱桃,樱桃依据花梗和花萼筒与前三者区分开。
3.3 花器官形态的演化
李朝銮[25]基于植物叶表皮解剖结构提出蔷薇科植物从温带类型向热带类型演化的趋势。黄文鑫等[7]基于叶脉序特征提出广义李属植物从温带类型向热带类型演化的趋势。曹菊逸[26]认为,花序中复合花序最原始,简单花序是由复合花序简化而成。蔷薇科花序演化的总方向是由复杂趋于简单,花朵的直径由小到大[16],由此反映出广义李属植物是从热带、亚热带类型向温带类型演化的趋势。从本文对花器官形态的研究可以看出,花器官微形态特征在属内各类群之间表现出连续性和过渡性,花序类型、花序梗长和花直径在广义李属各类群之间呈现出较为同步的演变。总状花序有稠李亚属、桂樱亚属、臀果木属和臭樱属,主要分布于中国的热带和亚热带地区;伞形或伞房状总状花序有樱亚属(除矮樱组),主要分布于中国的西南、华中和秦岭山脉的亚热带和温带地区;而花单生或2(3)朵簇生的李亚属、矮樱类、杏亚属和桃亚属多分布于中国北方的温带地区。从花器官形态上支持广义李属植物的演化从热带、亚热带类型向温带类型演化的趋势。
4 结 论
广义李属植物花器官形态具有一定共性和差异,其中花序类型、花序梗长、花直径以及总苞大小和形状可以作为属内的分类依据。本研究结果支持以下分类处理:桃属(AmygdalusLinn.)、杏属(ArmeniacaMill.)、李属(PrunusLinn.)、樱属、稠李属(PadusMill.)、桂樱属(LaurocerasusTourn. ex Duh.)和臭樱属归入广义李属内;矮樱类从樱亚属移至李亚属内,并作为李亚属下的矮樱组;崖樱桃和细花樱桃并入华中樱桃。
致谢:标本采集过程中得到中国科学院植物研究所张志耘研究员、华南农业大学俞新华老师、中山大学许可望博士、浙江省磐安县大盘山国家级自然保护区陈子林高级工程师和王盼工程师、福建省武夷山国家级自然保护区李冬静工程师以及辽宁省农业科学院果树科学研究所章秋平研究员等的帮助,在此一并表示感谢!
