确定遗传图距时重组值的校正机理分析
2021-07-01邢德智西安市临潼区代王初级中学陕西西安710611
邢德智 (西安市临潼区代王初级中学 陕西西安 710611)
三点测验(three-point test)是基因定位(gene mapping)最常用的方法,主要目的是根据基因之间的(染色体片段)交换值(cross-over value,交换百分率)确定基因之间的排列次序和相对距离进而绘制连锁遗传图(linkage map)。遗传图距(genetic distance)是由交换值确定的,但在三点测验中通过表现型(phenotype)数据测算出的直接结果是重组值(recombinant value,重组百分率)而非交换值;考虑到双交换和多交换,重组值理论上低于交换值,实际结果也往往反映出这样的关系;重组值(率)在任何情况下都不能大于0.5(50%)但是在作图函数中,与遗传距离对应的交换值可大于0.5。本文探讨两点测验和三点测验中交换值被低估的原因及其对应的校正方法,以及四点测验中交换值的计算方法及重组值的校正机理。
1 确定遗传图距时为什么要对重组值进行校正
1.1 重组值、交换值和遗传图距之间的关系 重组值和交换值的关系为:在2 个基因间遗传图距较短时,基因之间几乎只发生单交换,重组值接近交换值。遗传图距较远时,由于双交换、多交换发生机会增大,可检测出双交换的三点测验或重组值较大的两点测验中,重组值与交换值差别较大(详见1.3 和2.2),重组值和交换值的关系需要通过双交换、多交换率或作图函数进行校正。
根据定义,交换值与遗传图距之间的关系为:遗传图距是用交换值表示2 个基因之间的相对距离,以1%交换值去掉% 作为遗传图距单位(map unit,mu);为了纪念摩尔根(T. H. Morgen),该单位又称厘摩或分摩尔根(centi-Morgen,cM)。从测交实验数据计算出的是重组值而非交换值,在实际应用中往往需要将单交换重组值通过双交换重组值或作图函数校正为交换值。
1.2 三点测验中何时需1 倍、何时需2 倍双交换值校正 果蝇有刺眼(ec)、截翅(ct)和横脉缺失(cv)。将三杂合雌果蝇(ec ct+/+ +cv)与雄果蝇(ec ct cv/Y)测交,测交结果有8 种表现型,其中:亲组合型(ec ct+)为2 125 只、(+ +cv)为2 207只,单交换Ⅰ型(ec+cv)为273 只、(+ct+)为265只,单交换Ⅱ型(ec+ +)为217 只、(+ct cv)为223只,双交换型(+ + +)为5 只、(ec ct cv)为3 只;根据8 种 结 果 及 其 比 例 得 出ec、ct、cv为 连 锁 基 因而且顺序为ec-cv-ct,将测交结果整理为表1[1]进行连锁遗传分析。

表1 ec ct+/++cv×ec ct cv/Y测交结果[1]
分析表明,单交换Ⅰ重组值10.1%是ec-cv间发生单交换导致的,双交换重组值0.15%是ec-cv间和cv-ct间均发生1 次单交换导致的,但单交换Ⅰ重组值10.1%并未包括0.15%;同样道理单交换Ⅱ重组值8.3%也未包括0.15%。因此,三点测交实验中,在有双交换重组值时,如果用实测的单交换重组值直接替代相邻2 个基因间的交换值,则交换值一定被低估了,图距随之缩小了;当用2种实测的单交换重组值计算最远2 个基因间的交换值(18.4%)时,既未包括ec-cv间的0.15%也未包括cv-ct间的0.15%,自然要加上2 倍的双交换重组值才能接近最远2 个基因间的交换值。所以,用对应实测的单交换重组值加1 倍实测的双交换重组值计算ec-cv间和cv-ct间相邻基因间的交换值(10.1%+0.15%=10.3%;8.3%+0.15%=8.4%);用实测的单交换Ⅰ、Ⅱ重组值之和加2 倍的实测的双交换重组值计算ec-ct间最远基因间的交换值(10.1%+8.3%+2×0.15%=18.7%)。
这样才能正确地反映实际发生交换的频率。根据校正后的交换值绘制染色体图(chromosome map)(图1b)。如果不用2 倍双交换重组值校正2 种实测的单交换重组值,基因ec-cv间的图距是10.3,基因cv-ct间的图距是8.4 ,则基因ec-ct间的图距是18.4;不等于基因ec-cv和cv-ct之和18.7(图1a),所以,计算最远2 个基因间的交换值时,需要加上2 倍实测双交换重组值校正,其本质是分别校正了2 种实测的单交换重组值。

图1 果蝇基因ec-cv-ct 染色体图
1.3 为什么要用作图函数校正 当2 个连锁基因之间只发生单交换,重组值接近交换值,二者相当于一次函数关系。但当发生“二线双交换(或二线偶数次交换)”时,并不导致标记基因重组;发生“二线三交换(或二线奇数次交换)”时,标记基因重组的结果与单交换(一次交换)的结果完全相同;所以,对于交换导致重组的结果而言,偶数次交换相当于0 交换,除1 外的奇数次交换相当于单交换,即2 个连锁基因之间偶数次交换不导致重组只有奇数次交换才会导致重组[2];根据n个连锁基因性母细胞交换种类计算式C1n-1种单交换,C2n-1种双交换,C3n-1种三交换,…,Cn-1n-1种(n-1)交换,当n=2 时交换种类只有C12-1=1种单交换,即2 个连锁基因间不论发生多少次交换,测算出的只能是1 种“单交换”重组值,而这个“单交换”重组值完全没有反映出发生的偶数次交换、未完全反映出发生的除1 外的奇数次交换,从而导致这2 种情况下重组值都低于交换值,二者是非一次函数的特殊函数关系,自变量的变化值与函数的变化值不成正比例。因此,当2 个连锁基因间的距离较远重组值较大时,由于可能发生双交换或多交换(奇数次或偶数次),以及重组值中不反映(或不完全反映)2 个连锁基因之间发生了双交换或多交换,所以,重组值往往低于交换值,需要用霍尔丹(Haldane)作图函数校正(相关论述可参考遗传学教材[3])。
正因交换不一定导致重组,或交换与重组不是对应关系,重组值和交换值在这种情况下确实是不同的。
2 四点测验中交换值的计算方法及重组值的校正机理

2.1 四点测验中需要3 种双交换和1 种三交换重组值校正 假设“四点测验”有3 种单交换、3 种双交换、1 种三交换重组值,4 个基因在染色体上的顺序是a-b-d-e,单交换i、ii、iii 分别是a-b、b-d、d-e间各发生1 次交换导致的;双交换和三交换分别是单交换i、ii、iii 的i、ii,ii、iii,i、iii 或i、ii、iii同时发生导致的。各种交换的情况见表2 及图2,用3 段尺寸线表示在a-b、b-d、d-e间分别发生了1 次交换,染色体图下面表示染色体上同时发生了2 次交换且因交换位置不同而形成不同的双交换及同时发生了3 次交换形成三交换:

图2 基因a-b-d-e 交换图
由于四点测验的难度远远大于三点测验,而三点测验已解决了相关的实际问题,所以四点测验的资料较少见;表2 是一个四点测验的结果,则其交换值计算方法如下:
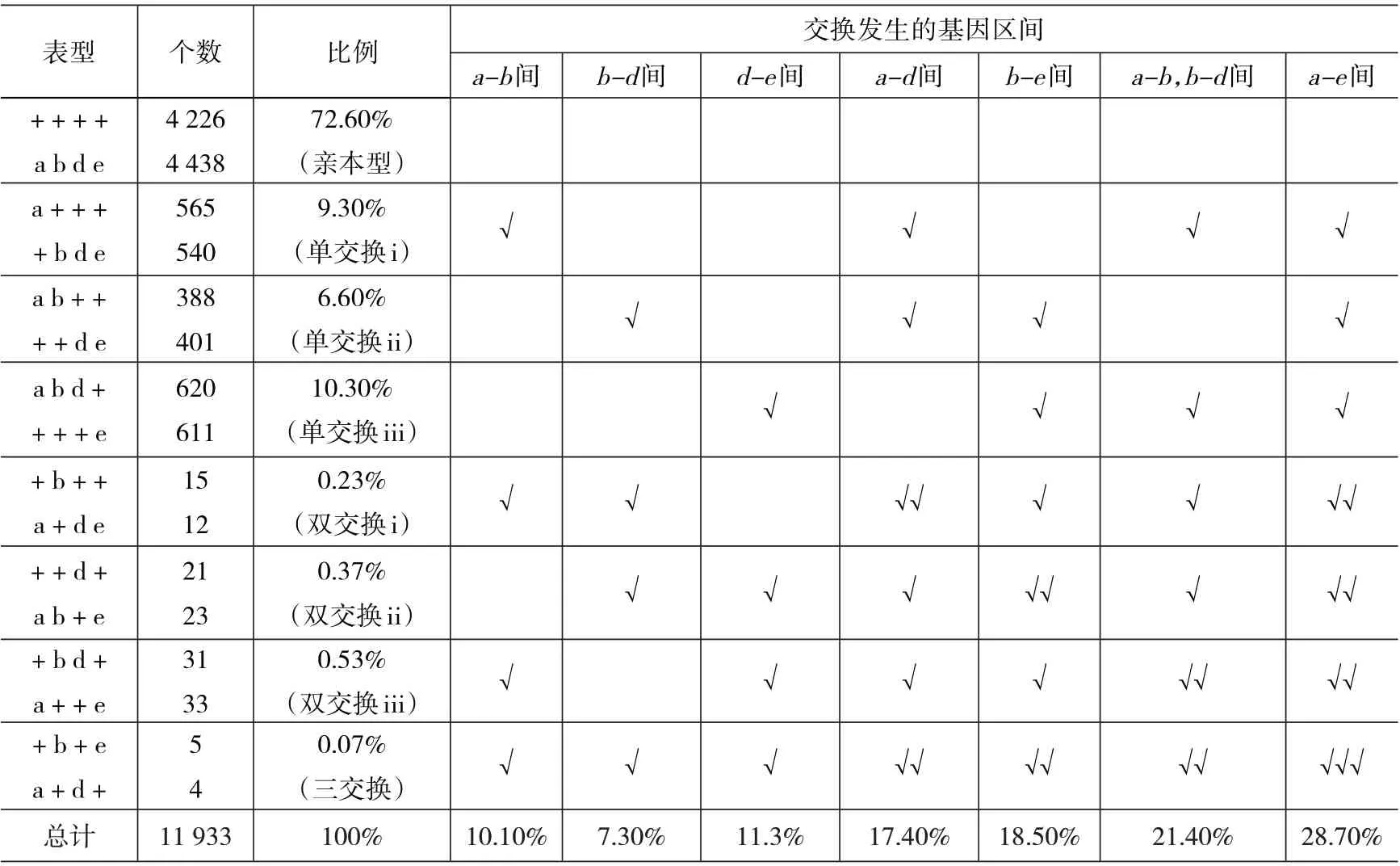
表2 四点测验结果分析
1)a-b、b-d、d-e相邻2 个基因1 段基因图距间交换值依次为:
①a-b间交换值=单交换i 重组值+1 倍(双交换i+双交换iii+三交换)重组值=9.3%+0.23%+0.53%+0.07%=10.1%。
②b-d间交换值=单交换ii 重组值+1 倍(双交换i+双交换ii+三交换)重组值=6.6%+0.23%+0.37%+0.07%=7.3%。
③d-e间交换值=单交换iii 重组值+1 倍(双交换ii+双交换iii+三交换)重组值=10.3%+0.37%+0.53%+0.07%=11.2%。
2)a-d、b-e相邻3 个基因2 段基因图距间交换值依次为:
①a-d间交换值=(单交换i+单交换ii)重组值+2 倍双交换i 重组值+1 倍(双交换ii+双交换iii)重组值+2 倍三交换重组值=9.3%+6.6%+2×0.23%+0.37%+0.53%+2×0.07%=17.4%。
②b-e间交换值=(单交换ii+单交换iii)重组值+1 倍 双 交 换i 重 组 值+2 倍 双 交 换ii 重 组 值+1 倍双交换iii 重组值+2 倍三交换重组值=6.6%+10.3%+0.23%+2×0.37%+0.53%+2×0.07%=18.5%。
3)a-e最远间4 个基因3 段基因图距交换值a-e间交换值=1 倍(单交换i+单交换ii+单交换iii)重组值+2 倍(双交换i+双交换ii+双交换iii)重组值+3 倍三交换重组值=9.3%+6.6%+10.3%+2×(0.23%+0.37%+0.53%)+3×0.07%=28.7%。
最远2 个基因之间的交换值28.7%与由a-b、b-d、d-e间交换值之和(10.1%+7.3%+11.2%)相吻合。反映出最远2 个基因之间的交换值=校正后所有连续相邻2 个基因间的交换值之和;即:最远2 个基因之间的遗传图距=所有连续相邻2 个基因间的遗传图距之和。
四点测验中交换值计算方法揭示出相邻2 个基因间、最远2 个基因间和任意2 个基因间的交换值计算规律是:
1)相邻2 个基因间的交换值=某1 种(对应的)单交换重组值+1 倍某2 种(相关)双交换重组值之和+1 倍三交换重组值;不论是几交换重组值,其对应的交换最多与相邻2 个基因间相关1次,计算时都用1 倍值。这间接说明两点测验检测不出2 个基因间发生了双交换和多交换,因此需要作图函数校正。
2)任意2 个基因间的交换值=相关的单交换重组值之和+相关双交换所涉及的交换次数×相关双交换的重组值+三交换所涉及的交换次数×三交换重组值;例如,b-e间的交换值=(单交换ii+单交换iii)的重组值+(1×双交换i 的重组值+2×双交换ii 的重组值+1×双交换iii 的重组值)+2×三交换的重组值,即6.6%+10.3%+0.23%+2×0.37%+0.53%+2×0.07%。此算式中交换值的计算规律更清楚地揭示出校正值“倍数”的本质是形成该重组配子时在所计算的2 个基因间的相关交换所涉及的交换次数。
3)表2 中每个区间的交换值等于该区间涉及的所有重组值之和;其中单交换重组值只用1 倍值,双交换和三交换重组值可能要用几倍值,用几倍值取决于它们在这个区间所涉及的交换次数(“√”的个数,无“√”空白处个数为0)。
“四点测验”中交换值的计算为什么能揭示出用几倍值校正的道理?“三点测验”中只有相邻2个基因1 段基因图距、相邻3 个基因2 段基因图距2 种交换值的计算,并且校正值只涉及1 种双交换值,因此根本无法说清楚“倍数”的本质。林毅在论述四点测验方法时,未说明四点测验中为什么产生3 种单交换、3 种双交换、1 种三交换,自然无法厘清单交换与双交换及三交换的关系;在计算交换值时只涉及4 个连锁基因中相邻2 个基因之间交换值的计算,同样无法说清楚“倍数”的本质。
本“四点测验”交换结果除了涉及7 种交换重组值外,还涉及相邻2 个基因1 段基因图距、相邻3 个基因2 段基因图距和最远4 个基因3 段基因图距3 种情况的交换值计算,它们都包括在“任意2个基因间交换值的计算”里。这个算式具有普遍意义并且校正时涉及多种双交换和三交换;而且用双交换值校正时情况较为复杂——例如,某种双交换与2 个基因间无关或相关1 次,最多相关2次,校正时对应的“倍数”分别为0、1、2。这样就可清楚地揭示校正值“倍数”的本质。
综上所述,多点测验中任意2 个基因间的交换值主要由相关的单交换重组值决定,但要估算出较准确的基因间交换频率,还需要用双交换、多交换重组值校正。用几倍重组值校正取决于它们在这2 个基因之间所涉及的交换次数。四点测验中不论是双交换重组值还是多交换重组值在相邻2 个基因间最多测出1 次交换所得的重组值,所以都是用1 倍值校正(a-b、b-d、d-e间交换值);双交换、多交换重组值是在2 段或3 段基因图距上发生2 次单交换及多次单交换导致的,所以通常用2 倍值、多倍值校正。总之,相邻2 个基因之间不论是几交换重组值,都是用1 倍值校正;最远2 个基因之间是几交换重组值,就用几倍值校正;任意2 个基因之间用几个双交换、多交换重组值校正取决于这2 个基因之间相关双交换、多交换的种类及种类个数,用几倍值校正取决于该交换在这2 个基因之间所涉及的交换次数。“四点测验”中任意2 个基因间双交换、三交换重组值校正时的“倍数”分别为0、1、2 和1、2、3,即双交换重组值最大用2 倍值进行校正,也可能用1 倍值或0 倍值去校正、三交换重组值最大用3 倍值校正,也可能用2 倍值或1 倍值进行校正。用0 倍值去校正指这2 个基因间的交换值与该种双交换重组值无关。
“三点测验”通常有2 种单交换,最多有1 种双交换,情况比“四点测验”简单。“三点测验”中因为形成双交换重组配子时,是中间基因与两侧相邻的2 个基因间分别交换1 次,即同时发生的2次交换也就是双交换,其重组的结果统计到实测双交换值里了,而相邻的2 个基因间的单交换重组值并未包括这部分双交换值,所以,估算相邻2个基因间的交换值,要用实测的单交换重组值加上1 倍双交换重组值校正;而形成双交换重组配子时在最远的2 个基因间(实际上为3 个基因2 段基因图距间)交换了2 次也未包括这部分双交换值,因此,估算最远2 个基因间的交换值,要用实测的2 种单交换重组值之和加上2 倍双交换重组值校正。有双交换重组值时,相邻2 个基因或最远2 个基因间的单交换重组值一定低于对应节段上基因间的交换值,只有通过1 倍或2 倍双交换重组值校正,才能正确地反映基因间实际发生交换的频率进而得到较为准确的遗传图距。例如,前述“果蝇有刺眼(ec)、截翅(ct)和横脉缺失(cv)”基因间交换值的计算。
这就是“三点测验”中交换值低估的原因,以及三点测验和四点测验中重组值校正的方法。
2.2 四点测验与Haldane 作图函数 两点测验无法检测出2 个基因间发生了双交换和多交换。如果四点测验中不考虑居间基因b和d,则无法反映出a-e间发生的3 种双交换和1 种三交换、只能检测出1 种单交换,其数值就是3 种单交换重组值之和(2 个标记基因间不论何处发生了1 次交换,结果形成同一类型的重组型配子);不校正之前交换值肯定被低估了。当考虑居间基因b和d,相当于在a-e间增添了2 个标记基因,就可检测出a-e间发生的双交换和多交换。
a-b、a-d和a-e间遗传图距依次增大,每一个区间涉及的交换总次数也相应增多(表2 中对应区间的对号总数,即4、8、12),这与Haldane 作图函数中,交换平均数越大,遗传图距也越大规律相吻合;或者说四点测验分析反映的规律,比三点测验更清楚地解释了Haldane 作图函数中,遗传图距随着交换平均数变化的道理。
3 小结
遗传图距是按交换值确定的,根据三点测交实验统计数据直接计算出的是重组值而非交换值,通过实测双交换重组值或作图函数校正间接计算出数值才是交换值。当有双交换重组值时,重组值一定小于交换值,一定要用双交换重组值校正,用几倍值校正取决于形成该种重组型配子时这2 个基因之间交换的次数;相邻2 个基因之间用1 倍值校正,最远2 个基因之间用2 倍值校正。“四点测验”中任意2 个基因间交换值计算式具有普遍意义,用重组值校正时涉及多种双交换和三交换,从而清楚地揭示出重组值校正值“倍数”的本质是相关交换在这2 个基因间所涉及的交换次数。当没有双交换重组值而且2 个连锁基因相距的遗传图距较远、重组值较大时,由于可能发生双交换或多交换,重组值也往往小于交换值,需要用Haldane 作图函数校正。
交换值确定遗传图距,它是个固定值。当2个基因的遗传图距较近只发生单交换时,交换值直接用单重组值替代;遗传图距较远发生双交换和多交换时,重组值要校正为交换值。所以,确定遗传图距时往往要对单交换重组值进行校正。本研究主要帮助学习者理解重组值的校正机理,减少交换值计算上的错误,加深对重组值、交换值概念及二者关系本质的理解,特别是对重组值校正“倍数”本质及Haldane 作图函数校正机理的理解。
