转录组测序分析毒死蜱与醚菊酯混配对二化螟毒杀的增效机制
2021-06-30徐鹿罗光华金瑜剑徐德进徐广春顾中言
徐鹿 罗光华 金瑜剑 徐德进 徐广春 顾中言
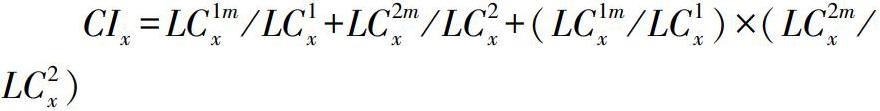
摘要: 为明确毒死蜱与醚菊酯混配对二化螟毒杀的增效机制,采用联合毒力指数法筛选毒死蜱与醚菊酯混配的增效比例,利用转录组测序分析毒死蜱与醚菊酯混配的增效机制。结果表明,毒死蜱与醚菊酯以1∶2(质量比)混配时表现出增效作用,Illumina HiSeqTM X Ten测序获得高质量的二化螟转录组。去离子水处理与毒死蜱处理、去离子水处理与醚菊酯处理、去离子水处理与增效混配剂处理、毒死蜱处理与增效混配剂处理、醚菊酯处理与增效混配剂处理的差异表达基因分别为465、392、1 874、117、25个。对毒死蜱与醚菊酯混配增效主要涉及的代谢过程进行通路分析,结合维恩交集法进行分析,发现解毒代谢酶可能介导毒死蜱与醚菊酯混配的增效机制。实时荧光定量PCR结果证实差异基因的表达量与测序数据匹配。本研究结果可为解析杀虫剂混配增效机制提供分子基础,为优化杀虫剂混配比例,建立二化螟抗性治理策略提供理论依据。
关键词: 二化螟;毒死蜱;醚菊酯;转录组测序;增效
中图分类号: S482.3;S481 文献标识码: A 文章编号: 1000-4440(2021)02-0317-09
Abstract: To reveal the synergistic mechanism of chlorpyrifos and ethofenprox mixture in poisoning Chilo suppressalis, the synergistic ratios of chlorpyrifos and ethofenprox mixture were screened by combined toxicity index method, the synergistic mechanism of chlorpyrifos and ethofenprox mixture were analyzed by transcriptome sequencing. The results indicated that chlorpyrifos and ethofenprox mixed at a 1∶2 mass ratio showed significant synergism on C. suppressalis. HiSeqTM X Ten platform was used to obtain C. suppressalis transcriptome with high quality data. 465, 392, 1 874, 117 and 25 differentially expressed genes were obtained between the groups of deionized water and chlorpyrifos, deionized water and ethofenprox, deionized water and synergistic mixture, chlorpyrifos and synergistic mixture, ethofenprox and synergistic mixture. Pathway analysis showed that synergistic effect of chlorpyrifos and ethofenprox mixture was mainly involved in the metabolic process, and it was speculated that detoxification metabolic enzymes might mediate synergism mechanism of chlorpyrifos and ethofenprox combined with Venn intersection algorithm. The expression of significant different genes was highly matched with transcriptome sequencing data by quantitative real-time polymerase chain reaction (PCR). The results can provide a molecular basis for analyzing synergism mechanism of insecticide mixtures, and provide a theoretical basis for optimizing ratio of insecticide mixtures and establishing resistance control strategy of C. suppressalis.
Key words: Chilo suppressalis;chlorpyrifos;ethofenprox;transcriptome sequencing;synergism
杀虫剂混配是杀虫剂应用的普遍技术,是治理害虫抗药性的重要策略[1]。害虫的抗药性会致使杀虫剂使用效率下降,导致可使用的杀虫剂数量减少。新型杀虫剂的研发速度不及害虫抗药性的发展速度。杀虫剂混配通过优化杀虫剂組合延缓害虫抗药性的发展[2]。杀虫剂混配存在增效、相加和拮抗3种作用,仅增效配比的杀虫剂可合理应用[3]。有研究发现,氯菊酯与毒死蜱增效混配杀虫剂可显著增加对抗性褐飞虱的毒力,提高田间的防治效果[4]。施用化学农药是治理害虫的主要措施,杀虫剂增效混配是治理害虫抗药性,延长杀虫剂使用寿命的重要手段,研究杀虫剂混配的增效机制是其应用的基础。杀虫剂混配增效作用是在害虫体内通过一种药剂影响另一种药剂或是2种药剂相互影响,从而提高混配杀虫剂的毒力[5]。有研究指出,杀虫剂混配产生增效是因为混配单剂之间相互影响,引起害虫体内部分生理生化变化,包括表皮穿透性的提高,害虫体内解毒代谢酶的抑制以及靶标部位作用能力增强3个方面[6],但目前对杀虫剂混配增效机制的认识仍然不足。转录组测序可用于研究害虫体内相关基因表达和调控路径,为解析杀虫剂混配增效机制提供新方法[7]。
二化螟[Chilo suppressalis(Walker)]是一种危害水稻生产的多食性害虫,主要分布于亚洲、中东和南欧,在中国长江流域及以南稻区危害严重。由于杂交稻的推广和耕作制度的改革,二化螟的发生量逐年增加,幼虫钻蛀取食水稻茎叶,造成严重减产[8]。据统计,中国二化螟年发生面积约为1.4×107 hm2,年经济损失约达1.15×1010元[9]。二化螟防治主要依靠化学农药,但二化螟对大多数常规杀虫剂已经产生了抗性[10]。毒死蜱在中国农业农村部取得登记,成为防治水稻害虫二化螟的常规药剂,但二化螟对毒死蜱已产生高水平抗性。醚菊酯是唯一投入到水稻田中使用的拟除虫菊酯类杀虫剂,二化螟对其处于敏感阶段[11]。毒死蜱作用于乙酰胆碱酯酶,而醚菊酯以钠离子通道为靶点,2种杀虫剂的作用方式不同,二化螟产生抗药性的机制不同,毒死蜱和醚菊酯可以混配使用。本研究拟筛选毒死蜱与醚菊酯混配的增效比例,通过转录组测序鉴定毒死蜱与醚菊酯混配的增效基因,解析混配杀虫剂的增效机制,以期为二化螟的治理以及农药减施增效提供理论依据。
1 材料与方法
1.1 供试虫源
本研究的供试虫源为2013年采自安徽省芜湖市水稻田的二化螟,在不接触任何杀虫剂的条件下,在室内以南粳46水稻连续饲养。饲养方法为无土水稻育苗法[10]。饲养条件为:温度(28±1) ℃,光周期16 L∶8 D,相对湿度>80%。
1.2 杀虫剂毒力测定
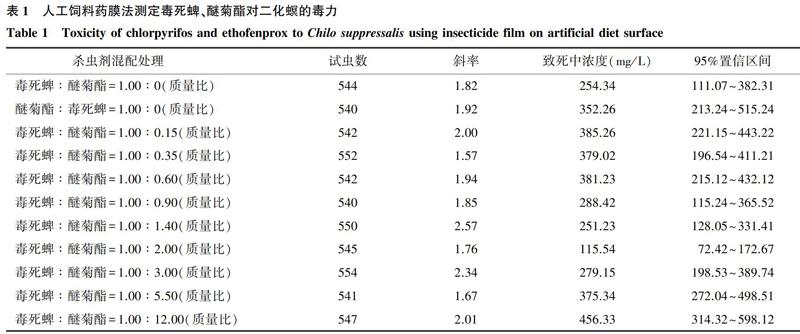
采用人工饲料药膜法[12]测定杀虫剂毒力。将97%毒死蜱(CPF)和95%醚菊酯(PEPE)原药用丙酮溶解于50 ml离心管中作为母液,用去离子水从中稀释出5~6个系列浓度,用来处理3龄二化螟幼虫,以去离子水处理为对照。采用Wang等[12]的方法制备人工饲料,在每孔饲料表面加入100 μl药液,置于室温下晾干。每孔接入1头3龄幼虫,每个浓度处理48头,重复3次,接虫后用黑色棉布覆盖,再盖上培养皿盖,并用橡皮筋扎紧,饲养72 h后检查死亡情况,统计死亡数并计算致死中浓度(LC50)。
1.3 杀虫剂联合作用测定
采用联合毒力指数(CI)法评估毒死蜱与醚菊酯混配的增效、拮抗、相加作用[13],通过毒死蜱和醚菊酯的LC50设置混配比例,CI的计算公式为:
LC1mx和LC2mx分别指当死亡率为x时杀虫剂1和杀虫剂2混用的致死剂量;LC1x和LC2x分别指死亡率为x时杀虫剂1和杀虫剂2单独使用的致死剂量;CIx是当死亡率为x时的联合毒力指数。当杀虫剂混合使用表现为相加作用时,CI=1;当杀虫剂混合使用表现为增效作用时,CI<1;杀虫剂混合使用表现为拮抗作用时,CI>1。死亡率为50%的情况下,计算混配杀虫剂的毒力及其联合作用的CI值。混配杀虫剂的毒力用混合使用时和单独使用时的死亡率差值计算,其增效作用的毒力计算公式为:M(1+2)=100%-(M1+M2)。M1和M2:分別指杀虫剂1和杀虫剂2单独使用时的死亡率;M(1+2):指杀虫剂1和杀虫剂2混配使用时的死亡率。
1.4 转录组测序
使用毒死蜱、醚菊酯以及毒死蜱与醚菊酯混配剂处理30~40头二化螟3龄幼虫6 h,每个处理重复3次,去离子水处理作为对照,处理后样品放入液氮中冷冻,-80 ℃保存备用。采用Trizol试剂盒提取二化螟总RNA,用1%琼脂糖凝胶电泳和核酸蛋白定量仪检测提取的总RNA的完整度、纯度和浓度,-80 ℃保存备用。样品cDNA文库的构建和测序由上海祥音生物科技有限公司操作,测序在Illumina HiSeqTM X Ten仪器上完成,重复3次。将测序的原始数据去除adaptor序列和低质量的读长得到过滤数据,用HISAT v2.0.4软件比对二化螟基因组进行分析,使用TopHat v2.0.11软件比对参考基因组。用Bowtie2 v2.2.6软件分析每百万基因片段中来自某一基因每千碱基长度的片段数目,评估基因表达丰度(FPKM),利用DEGSeq软件分析2个样本之间的差异表达基因,采用维恩作图软件v1.6.17分析多个样本之间的共表达差异基因。
1.5 实时荧光定量PCR验证
用SV Total RNA Isolation System Kit试剂盒提取二化螟幼虫总RNA,按方法1.4的方法对总RNA的完整度、纯度和浓度进行检测并保存。cDNA第一链的合成采用PrimeScriptTM RT Reagent Kit试剂盒,按说明书操作。采用荧光定量PCR(qRT-PCR)技术在ABI Prism 7300定量PCR仪上验证鉴定的毒死蜱与醚菊酯混配增效基因表达量变化。混配增效基因和内参基因(Actin A1和G3PDH)引物使用Beacon Designer 7.0软件进行设计,由生工生物工程(上海)股份有限公司合成,根据SYBR Green Premix Ex Taq的试剂盒说明书进行扩增。采用PCR仪自带软件进行数据分析,设3个生物学重复。采用2-△△Ct法计算目标基因的相对表达量。
1.6 数据处理
用Abbott公式校正各处理死亡率。利用POLO Plus逻辑几率值软件中的几率值分析评估死亡率为50%的药剂剂量、置信区间以及几率死亡率与剂量对数的斜率。采用单因素方差分析中的Tukey显著性差异检测进行多组比较。所有统计分析均采用SPSS13.0软件进行。
2 结果与分析
2.1 毒死蜱与醚菊酯混配对二化螟毒杀的增效、拮抗作用
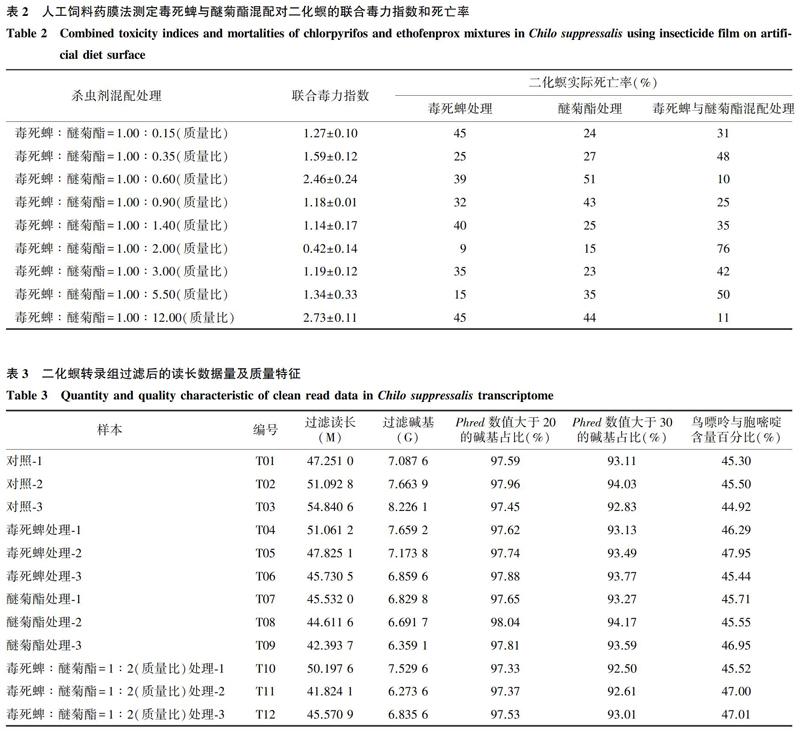
毒死蜱、醚菊酯对二化螟3龄幼虫的LC50分别为254.34 mg/L和352.26 mg/L(表1)。毒死蜱与醚菊酯以1∶2(质量比)混配,致死中浓度为115.54 mg/L,联合毒力指数为0.42(表2),表现出增效作用。
2.2 二化螟转录组特征分析
通过Illumina HiSeqTM X Ten平台对毒死蜱、醚菊酯、毒死蜱与醚菊酯增效混配剂、去离子水处理构建的12个cDNA文库(T01~T12)进行测序,4个处理分别产生48.205 6 M、44.179 1 M、45.864 2 M和51.061 5 M的平均过滤读长,产生7.230 9 G、6.626 9 G、6.879 6 G和7.659 2 G的平均过滤碱基,测序质量控制后显示Phred数值大于20和30的碱基占比分别在97.00%和90.00%以上。所有cDNA文库的鸟嘌呤与胞嘧啶含量百分比为44.92%~47.95%,获得了高质量的二化螟转录组(表3)。二化螟转录组的原始数据提交到NCBI,获取短读存档登录号(S ̄R ̄R ̄1 ̄1 ̄4 ̄9 ̄6 ̄7 ̄2 ̄0 ̄、S ̄R ̄R ̄1 ̄1 ̄5 ̄0 ̄8 ̄5 ̄1 ̄9 ̄、S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄2 ̄5 ̄6 ̄4 ̄、S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄3 ̄3 ̄9 ̄9 ̄、 ̄S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄2 ̄5 ̄9 ̄8 ̄、 ̄SRR11612564、SRR11612730、SRR1161283 ̄5 ̄、 ̄S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄2 ̄9 ̄3 ̄8 ̄、S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄3 ̄3 ̄5 ̄9 ̄、S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄3 ̄3 ̄6 ̄0和S ̄R ̄R ̄1 ̄1 ̄6 ̄1 ̄3 ̄3 ̄9 ̄6 ̄)。
2.3 毒死蜱与醚菊酯混配增效处理的差异基因表达量及通路分析
基于FPKM分析毒死蜱、醚菊酯、毒死蜱与醚菊酯增效混配剂、去离子水处理下样品的相关性,各处理下样品间基因表达的皮尔逊相关系数分别为0.82、0.91、0.94和0.98(图1),表明获得的测序数据可靠。去离子水处理与毒死蜱处理、去离子水处理与醚菊酯处理、去离子水处理与增效混配剂处理、毒死蜱处理与增效混配剂处理、醚菊酯处理与增效混配剂处理的差异表达基因(DEG)(图2A)分别为465、392、1 874、117、25个,其表达量比值分别为-3.91~4.38、-4.11~6.25、-5.66~10.95、-2.65~3.24和-2.01~2.48,样品间基因表达模式存在显著差异(图2B)。采用京都基因与基因组百科全书(KEGG)通路富集分析DEG的功能,去离子水处理与毒死蜱处理之间的465个DEG主要富集代谢中的全局代谢总括(41个)、脂质代谢(28个)、外源物降解代谢(23个)以及富集环境信息处理的信号转导(27个)(图3A);去离子水处理与醚菊酯处理之间的392个DEG主要富集代谢中的全局代谢总括(61个)、脂质代谢(33个)、外源物降解代谢(24个)以及富集环境信息处理的信号转导(20个)(图3B);去离子水处理与增效混配剂处理之间的1 874个DEG主要富集代谢中的全局代谢总括(232个)、核苷酸代谢(127个)、碳水化合物代谢(90个)、氨基酸代谢(84个)、脂质代谢(82个)、外源物降解代谢(59个),富集环境信息处理的信号转导(140个)、信号分子与相互作用(78个)、膜传输(65个),富集遗传信息处理的转录(70个)、翻译(54个)以及折叠、分类和降解(52个),富集细胞过程的细胞群落(58个)以及细胞生长和死亡(73个)(图3C);毒死蜱处理与增效混配剂处理之间的117个DEG主要富集代谢中的全局代谢总括(13个)、核苷酸代谢(6个)、碳水化合物代谢(6个)、氨基酸代谢(7个)、脂质代谢(8个)、外源物降解代谢(6个)以及富集环境信息处理的信号转导(11个)(图3D);醚菊酯处理与增效混配剂处理之间的25个DEG主要富集代谢中的全局代谢总括(6个)、碳水化合物代谢(2个)、氨基酸代谢(2个)、脂质代谢(2个)、外源物降解代谢(2个)以及富集环境信息处理的信号转导(2个)(图3E)。上述研究结果表明,毒死蜱与醚菊酯混配增效机制主要涉及代谢过程。结合维恩交集法筛选去离子水处理与增效混配剂处理、毒死蜱处理与增效混配剂处理、醚菊酯处理与增效混配剂处理三者的共表达差异基因,结果(图4A)发现CYP341B2、同向转运体和核酸酶基因HARBI1共表達,代谢通路中的解毒代谢酶可能介导毒死蜱与醚菊酯的增效机制。
采用qRT-PCR分析上调的CYP341B2、UGT340C2、CYP341A15、气味结合蛋白70基因(OBP70)、多抗药相关蛋白基因(MRP)、表皮蛋白22基因(CP22)、表皮蛋白27基因(CP27),以及下调的热激蛋白90基因(Hsp90)、CYP6AB45、CYP6AE60的表达量,结果(图4B)表明qRT-PCR基因表达量与转录组测序数据相匹配。
3 讨论
杀虫剂轮用和混配是治理害虫抗药性的普遍策略,在单一杀虫剂使用模式下,轮用杀虫剂并不能杀死全部抗性害虫,而在杀虫剂混用模式下,理论上可以杀死全部抗性害虫[14]。有研究结果表明,二化螟对敌百虫、对硫磷、甲基对硫磷、硫磷、杀虫单、杀虫双、氟虫腈(已禁用)、阿维菌素和双酰胺类杀虫剂已经产生了中等至高等水平的抗性[15-17]。二化螟幼虫于水稻茎秆中渡过生命周期,这层天然屏障加大了防治难度,但施用化学农药仍是治理二化螟的重要手段。由于二化螟具有较强的抗药性,可以考虑将无交互抗性的敏感药剂与抗性药剂混用。毒死蜱与醚菊酯以1∶2(质量比)混配明显增强对二化螟的毒力,混配剂的致死效果优于单剂,可以延长化学农药的使用寿命。要想优化毒死蜱与醚菊酯的混配比例,促进杀虫剂持续使用,需解析毒死蜱与醚菊酯混配剂的增效机制。本研究利用转录组测序方法分析毒死蜱与醚菊酯的协同增效机制,转录组测序产生44.179 1~51.061 5 M的平均过滤读长和6.626 9~7.659 2 G的平均过滤碱基,高于报道的二化螟转录组数据[18],检测到的表达基因总数与二化螟基因组中注释的基因总数接近[19],获得的Phred数值大于20和30的碱基数占比以及鸟嘌呤与胞嘧啶含量百分比数据,反映出高的测序质量。皮尔逊相关系数是检测试验可靠性和样本选择的重要指标,二化螟生物学重复样品间皮尔逊相关系数大于0.8,表明样品间的表达模式相似度高,确保后续的差异基因分析。
DEG的KEGG通路富集分析结果显示,毒死蜱与醚菊酯增效混配差异表达基因主要分布于代谢、环境信息处理、遗传信息处理和细胞过程,主要参与碳水化合物代谢、氨基酸代谢、脂质代谢、信号转导、外源物降解代谢。碳水化合物代谢作为生物体内重要的代谢途径,为细胞中许多生理反应和过程提供能量和底物来源。毒死蜱与醚菊酯增效混配剂处理二化螟后碳水化合物代谢基因数量明显增加,表明二化螟解毒毒死蜱与醚菊酯增效混配剂会消耗大量能量。氨基酸代谢通过氧化产生能量满足机体需要,氨基酸参与杀虫剂的代谢过程。二化螟暴露于毒死蜱与醚菊酯增效混配剂中,氨基酸代谢通路中多个基因显著表达,表明毒死蜱与醚菊酯混配剂的T01~T12见表3。
协同增效过程需要氨基酸的合成和分解。当糖缺乏时,脂质代谢可为生物体提供能量,是细胞内重要的生理过程。脂质代谢通路中的多个基因在二化螟暴露于毒死蜱与醚菊酯增效混配剂中后显著表达,推测这些差异表达基因可能会改变脂质代谢,从而影响二化螟的繁殖。信号转导涉及生物体的各种生理过程[20-21],二化螟暴露于毒死蜱与醚菊酯增效混配剂中后,差异表达基因数量高于其他通路,推断二化螟接触杀虫剂后通过信号转导通路传递杀虫剂信号,进而启动各种生理反应避免杀虫剂致死效应。外源物降解代谢是昆虫接触杀虫剂后产生的主要生理过程,解毒代谢酶和转运体参与其中,解毒代谢过程是一个生化代谢途径,将外源物分解成无毒的形式,本研究鉴定出解毒代谢差异共表达基因,表明解毒代谢酶可能介导毒死蜱与醚菊酯混配剂协同增效机制。本研究鉴定的代谢过程相关通路的差异表达基因在毒死蜱与醚菊酯混配剂对二化螟毒杀增效中起到关键作用,但其功能还需进一步研究,昆虫杆状病毒表达系统和CRISPR/Cas9基因编辑可为其功能研究提供支持[22-23]。本研究结果表明,毒死蜱与醚菊酯在特定比例下混配对二化螟毒杀产生增效作用,通过转录组测序分析毒死蜱与醚菊酯对二化螟毒杀的增效机制,其结果为杀虫剂混配剂配比的优化以及二化螟抗性治理策略的建立提供理论依据。
参考文献:
[1] 顾中言,许小龙,徐德进,等. 杀虫剂混合使用的增效作用评判分析[J]. 农药学学报,2009,11(3):304-311.
[2] TONG S M, FENG M G. A mixture of putative sodium salts of camptothecin and bamboo tar is a novel botanical insecticide against rice planthoppers and stem borers [J]. Journal of Pest Science, 2016, 89: 1003-1011.
[3] PERRE C D, MURPHY T M, LYDY M J. Mixture toxicity of phostebupirim and cyfluthrin: species-specific responses [J]. Environmental Toxicology and Chemistry, 2017, 36(7): 1947-1954.
[4] YU Y Y, TING L C. Synergistic effect and field control efficacy of the binary mixture of permethrin and chlorpyrifos to brown planthopper (Nilaparvata lugens) [J]. Journal of Asia-Pacific Entomology, 2019, 22(1): 67-76.
[5] ATTIQUE M, KHALIQ A, SAYYED A. Could resistance to insecticides in Plutella xylostella (Lep., Plutellidae) be overcome by insecticide mixtures?[J]. Journal of Applied Entomology, 2006, 130(2): 122-127.
[6] 余海明,魏方林,朱國念. 三唑磷与氟虫腈复配对粘虫的增效机理研究[J]. 农药学学报,2009,11(3):317-322.
[7] XU L, LUO G H, SUN Y, et al. Multiple down-regulated cytochrome P450 monooxygenase genes contributed to synergistic interaction between chlorpyrifos and imidacloprid against Nilaparvata lugens [J]. Journal of Asia Pacific Entomology, 2020, 23: 44-50.
[8] JIANG W H, JIANG X J, YE J R, et al. Rice striped stem borer, Chilo suppressalis (Lepidoptera: Pyralidae), overwintering in super rice and its control using cultivation techniques [J]. Crop Protection, 2011, 30(2): 130-133.
[9] 姜玉英,刘万才,黄 冲,等. 2020年全国农作物重大病虫害发生趋势预报[J]. 中国植保导刊,2020,40(2):37-53.
[10]XU L, ZHAO J, SUN Y, et al. Constitutive overexpression of cytochrome P450 monooxygenase genes contributes to chlorantraniliprole resistance in Chilo suppressalis (Walker) [J]. Pest Management Science, 2019,75(3):718-725.
[11]韩 平. 二化螟对醚菊酯的抗性及相关代谢酶基因的研究[D]. 南京:南京农业大学, 2014.
[12]WANG B J, WANG Y, ZHANG Y, et al. Genome-wide analysis of esterase-like genes in the striped rice stem borer, Chilo suppressalis [J]. Genome, 2015, 58(6): 323-331.
[13]CHOU T C, TALALAY P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors [J]. Advances in Enzyme Regulation, 1984, 22: 27-55.
[14]SHI X Q, XIONG M H, JIANG W H, et al. Efficacy of endosulfan and fipronil and joint toxic action of endosulfan mixtures against Leptinotarsa decemlineata (Say) [J]. Journal of Pest Science, 2012, 85: 519-526.
[15]姜衛华. 二化螟的抗药性及综合防治研究[D]. 南京:南京农业大学, 2011.
[16]YAO R, ZHAO D D, ZHANG S, et al. Monitoring and mechanisms of insecticide resistance in Chilo suppressalis (Lepidoptera: Crambidae), with special reference to diamides [J]. Pest Management Science, 2017, 73: 1169-1178.
[17]LI X X, LI R, ZHU B, et al. Overexpression of cytochrome P450 CYP6BG1 may contribute to chlorantraniliprole resistance in Plutella xylostella (L.) [J]. Pest Management Science, 2018, 74(6): 1386-1393.
[18]MENG X, DONG F, KUN Q, et al. Transcriptome analysis reveals global gene expression changes of Chilo suppressalis in response to sublethal dose of chlorantraniliprole [J]. Chemosphere, 2019, 234: 648-657.
[19]MA W H, ZHAO X X, YIN C L, et al. A chromosome level genome assembly reveals the genetic basis of cold tolerance in a notorious rice insect pest, Chilo suppressalis [J]. Molecular Ecology Resources, 2020, 20: 268-282.
[20]周大颜,张志新,黄彩林,等. 3种白鲫杂交子代的转录组学分析[J]. 南方农业学报,2019,50(6):1328-1338.
[21]潘宝贵,钱恒彦,戈 伟,等.辣椒应答冷信号转导机制研究进展[J].江苏农业学报,2019,35(3):743-748.
[22]ELZAKI M E A, MIAH M A, PENG Y C, et al. Deltamethrin is metabolized by CYP6FU1, a cytochrome P450 associated with pyrethroid resistance in Laodelphax striatellus [J]. Pest Management Science, 2018, 74: 1265-1271.
[23]WANG H D, SHI Y, WANG L, et al. CYP6AE gene cluster knockout in Helicoverpa armigera reveals role in detoxification of phytochemicals and insecticides [J]. Nature Communications, 2018, 9: 4820.
(责任编辑:王 妮)
