小熊猫源犬瘟热病毒株H、F基因的克隆及序列分析
2021-06-30翟俊琼卜婉迪谢逸伦刘灿彬罗满林
蒋 梅,陈 武,翟俊琼,卜婉迪,谢逸伦,刘灿彬,单 芬*,罗满林*
(1.华南农业大学兽医学院,广州 510642; 2.广州动物园 广州市野生动物研究中心,广州 510070)
犬瘟热(cannine distermper, CD)由犬瘟热病毒(canine distermper virus, CDV)感染引起的高度接触性、致病性传染病,呈世界范围内广泛分布[1]。临床症状主要有发热、咳嗽、腹泻、呼吸困难、呕吐甚至死亡[2-3]。CDV是一种有包膜、不分节段、无重叠的单链负股RNA病毒,属于副黏病毒科(Paramyxoviride),麻疹病毒属(Morbillivirus)[4]。病毒基因组全长为15 690 bp,从3′到5′分别编码核衣壳(N)、磷蛋白(P)(包含两个非结构蛋白基因,C和V)、基质蛋白(M)、融合蛋白(F)、血凝素蛋白(H)、大蛋白(L)8个蛋白[5]。H和F糖蛋白是病毒粒子诱导中和保护性抗体的最外层蛋白质,比其他CDV蛋白更易变异,并决定病毒感染的宿主特异性[6]。两种糖蛋白都是诱导针对CDV的保护性免疫反应的抗原决定簇,H和F基因的遗传变异被认为是宿主感染CDV数量增加的重要原因,因此对CDV毒株H、F基因进行深入研究更为重要[7-8]。
小熊猫(Ailurusfulgens),国家二级保护动物,目前多被圈养于动物园或野生动物救护站[9]。犬瘟热病毒已经成为我国圈养小熊猫大量死亡、种群数量急剧下降的主要病原,常与犬冠状病毒、犬细小病毒发生混合感染,危害严重[10-11]。1968年,米克维茨[12]首次报道犬瘟热病毒导致小熊猫的大规模疾病和死亡,随后,中国多地相继报道圈养小熊猫感染犬瘟热病毒的病例,包括因接种犬瘟热弱毒疫苗导致死亡的案例等[13]。本研究中的毒株来自江苏某地封闭管养圈养的小熊猫,此前从未发生过犬瘟热感染,所以,探究病毒的遗传演化背景,了解病毒基因的变异情况对疾病防控具有指导意义。
1 材料与方法
1.1 材料
1.1.1 病料、菌株 病料采集自江苏某地感染犬瘟热病毒病死小熊猫的肺组织;大肠杆菌DH5α感受态购自天根生化(北京)科技有限公司。
1.1.2 主要试剂 病毒基因组DNA/RNA提取试剂盒,购自AXYGEN生物技术有限公司产品;胶回收试剂盒,购自OMEGA公司产品;DNA Marker、pMD18-T 载体,PrimerScriptTMOne step RT-PCR Kit,均购自宝生物(大连)有限公司;氨苄青霉素(Amp+)、LB培养基(酵母提取物2 g、胰蛋白胨1 g和氯化钠2 g,定容至200 mL)均购自广州华奇盛生物科技有限公司。
1.2 方法
1.2.1 病料处理 病料组织置于4倍体积的PBS中进行充分研磨,分装后4 ℃ 5 000 r·min-1离心10 min,弃沉淀,收取的上清液用0.22 μm滤器过滤除菌,保存于-80 ℃备用。
1.2.2 基因克隆及测序 比较分析GenBank上发表的犬瘟热病毒N基因序列,设计合成用于鉴定感染CDV的特异性鉴定引物,比较分析GenBank上发表的H、F基因全长序列,设计扩增H及F基因的引物,引物由北京睿博兴科生物技术有限公司合成,引物详细情况见表1。扩增的PCR产物经琼脂糖凝胶回收试剂盒回收后,与pMD18-T 载体连接,连接产物转化至大肠杆菌DH5α 感受态细胞,初步鉴定正确的重组质粒送生工生物工程(上海)股份有限公司测序。

表1 扩增CDV的引物设计
1.2.3 序列分析 采用Lasergene Version 7.1 软件中SeqMan、Meglign的对序列进行拼接、序列比对、抗原表位及分子特征分析。测得的序列登陆NCBI进行BLAST分析。同时下载不同基因型的犬瘟热病毒代表序列进行基因、氨基酸序列相似性分析。采用MEGA7.1软件绘制H、F基因的系统进化树。进化树的绘制方法:应用Neighbor-joining法(参数设置为1 000 replications)及Maximum composite likelihood model 比对核苷酸序列。采用NetNGlyc1.0软件进行糖基化位点分析。
2 结 果
2.1 鉴定引物RT-PCR扩增及H、F全长基因扩增
采用针对CDVN基因的特异性鉴定引物经RT-PCR 扩增、1%琼脂糖凝胶电泳之后,得到大小约为637 bp的目的基因片段,结果与预期片段大小一致(图略),将PCR产物送生工生物工程(上海)股份有限公司进行序列测定,比对分析证实为CDVN基因序列。采用针对H、F基因全长的特异性引物,对H、F的全长基因进行扩增,产物经1%琼脂糖凝胶检测,获得与预期大小一致的H基因全长片段(1 824 bp)和F基因全长片段(1 989 bp),结果如图1所示。
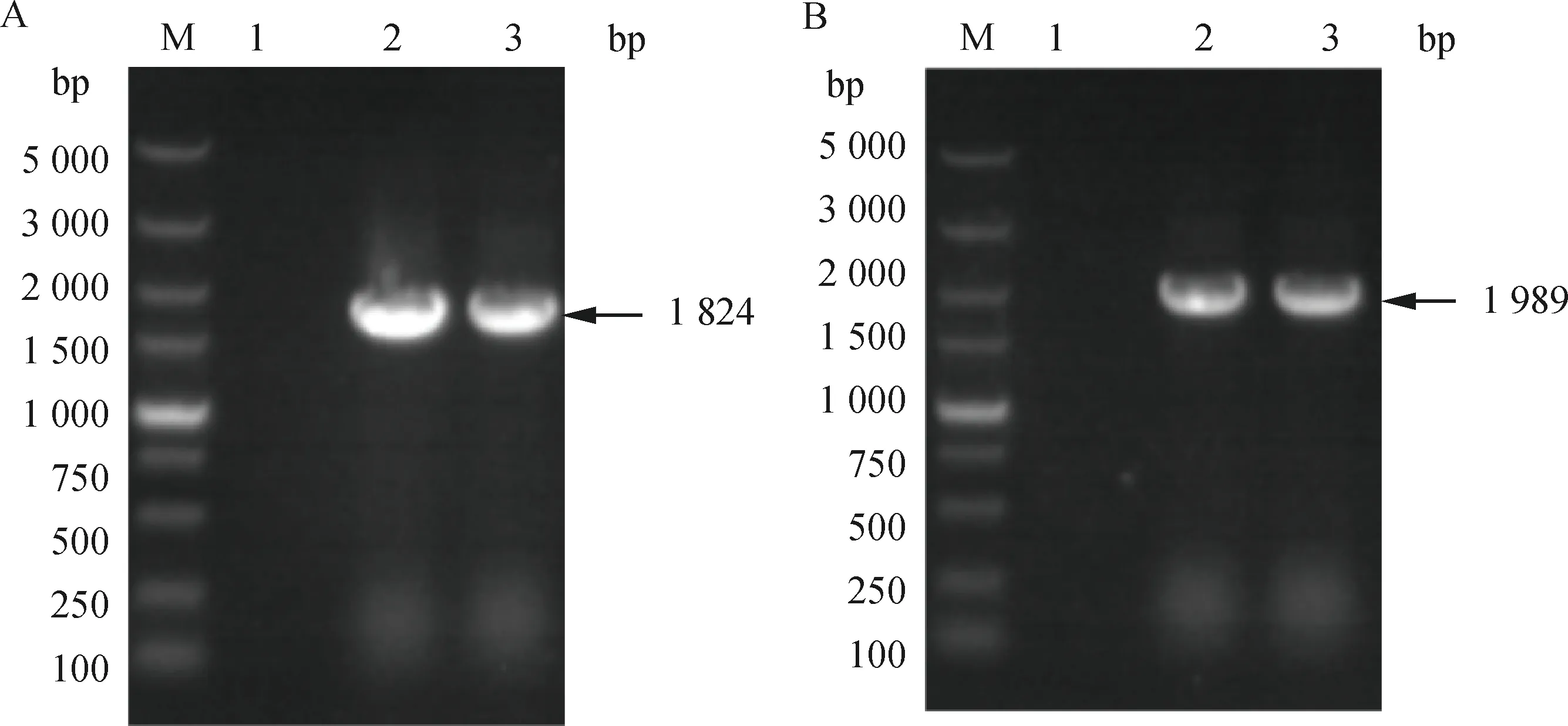
A. GD-1 H基因扩增;B. GD-1 F基因扩增;M. DL5000 DNA相对分子质量标准;1. 阴性对照;2. 扩增片段;3. 阳性对照
2.2 H、F基因的序列及遗传演化分析
H基因序列测定结果表明:H基因全长为1 824 bp, 编码607个氨基酸。登录GenBank进行Blast分析,H基因序列与丹麦报道的登录号为GU266280犬源犬瘟热病毒株的核苷酸序列相似性最高,为96%。与其他参考毒株的核苷酸相似性为88.5%~95.4%,相应的氨基酸序列相似性为83.1%~95.2%,与标准强毒株A75/17(登录号:AF164967,犬源CDV)氨基酸相似性为95.2%;与Convac、Onderstepoort、Recombinant Snyder Hill、CDV3四种经典疫苗株同源性较低,核苷酸相似性为90.3%~90.8%,对应的氨基酸序列相似性为88.2%~89.3%。
F基因测序结果表明,F基因全长为1 989 bp,编码663个氨基酸,其中1—135位是信号肽,136—224位为F2,225—662位为F1,切割位点的氨基酸序列为A↓QIHW。登录GenBank进行Blast分析,与巴西报道的登录号KY057355犬源犬瘟热病毒的核苷酸相似性最高,为95.7%;与其他CDV毒株相应的基因序列核苷酸相似性为92.2%~95.6%,其推定的氨基酸相似性为91.1%~99.0%,与疫苗株Onderstepoort、Recombinant Snyder Hill、CDV3、Shusiky等相应序列的核苷酸、氨基酸序列相似性较低,分别为91.0%~91.2%和89.1%~89.7%。
应用MEGA 7.0软件将GD-1株H、F基因与Asia-1、Asia-2、Asia-3、Asia-4、America-1、America-2、Europe、Europe-wildlife、Rockborn-like和Arctic-like等10个基因型的参考毒株进行遗传演化分析。结果发现:GD-1株分离株与Asia-4型的各参考毒株亲缘关系较近,位于同一大的进化分支,但是单独形成一个小的亚基因型分支,而与疫苗株America-1 亲缘关系远,这与国内报道的流行毒株位于Asia-1型存在差异(图2)。
2.3 H、F基因的分子特征分析
对H、F蛋白基因进行N-糖基化位点分析,结果显示:H蛋白基因具有8个N-糖基化位点,分别在19、149、309、391、422、456、587、603位点。其中,309—311糖基化位点是野毒株所特有的;其他位点没有发现新增的糖基化位点,这和大量文献中报道的野毒株含有8个或9个糖基化位点相一致;F蛋白共有6个N-糖基化位点,分别位于62、108、141、173、179、517位。
对H、F基因编码氨基酸序列进行蛋白抗原表位预测,H蛋白的抗原表位在1—38、368—391、419—433、568—607位氨基酸,与CDV3、Convac、recombinant Snyder Hill和Onderstepoort 疫苗株存在差异,与疫苗株Convac株、Onderstepoort株在393—408位氨基酸的抗原表位也有不同,与经典野毒株A75/17比对,在419—450、568—607位氨基酸处有较大差异。对F基因编码的F0蛋白进行抗原表位预测比对分析,在1—133、176—194、629—663位氨基酸的抗原表位与疫苗株(CDV3株、Shusiky株、recombinant Snyder Hill株和Onderstepoort株)和经典野毒株A75/17相应区域差异明显。
CDV H蛋白作为重要的膜蛋白,主要通过与宿主体内SLAM受体结合而复制。研究表明H蛋白个别氨基酸位点改变会影响与SLAM受体的结合能力。文献报道这些位点集中在525、526、529、530、549等,分析发现本分离株的这几个位点氨基酸与欧亚野生毒株的完全一致,与疫苗株相比530、549位氨基酸差异明显。
此外CDV H及F蛋白单个氨基酸的变异,可能引起毒力发生改变。分析发现该毒株的H蛋白氨基酸序列与其他CDV参考毒株相比,在24、41、85、219等共计9个氨基酸位点发生明显变异(表2)。与其他参考毒株相比, F蛋白共计有115、130、162等11处氨基酸发生了新的变异(表2)。
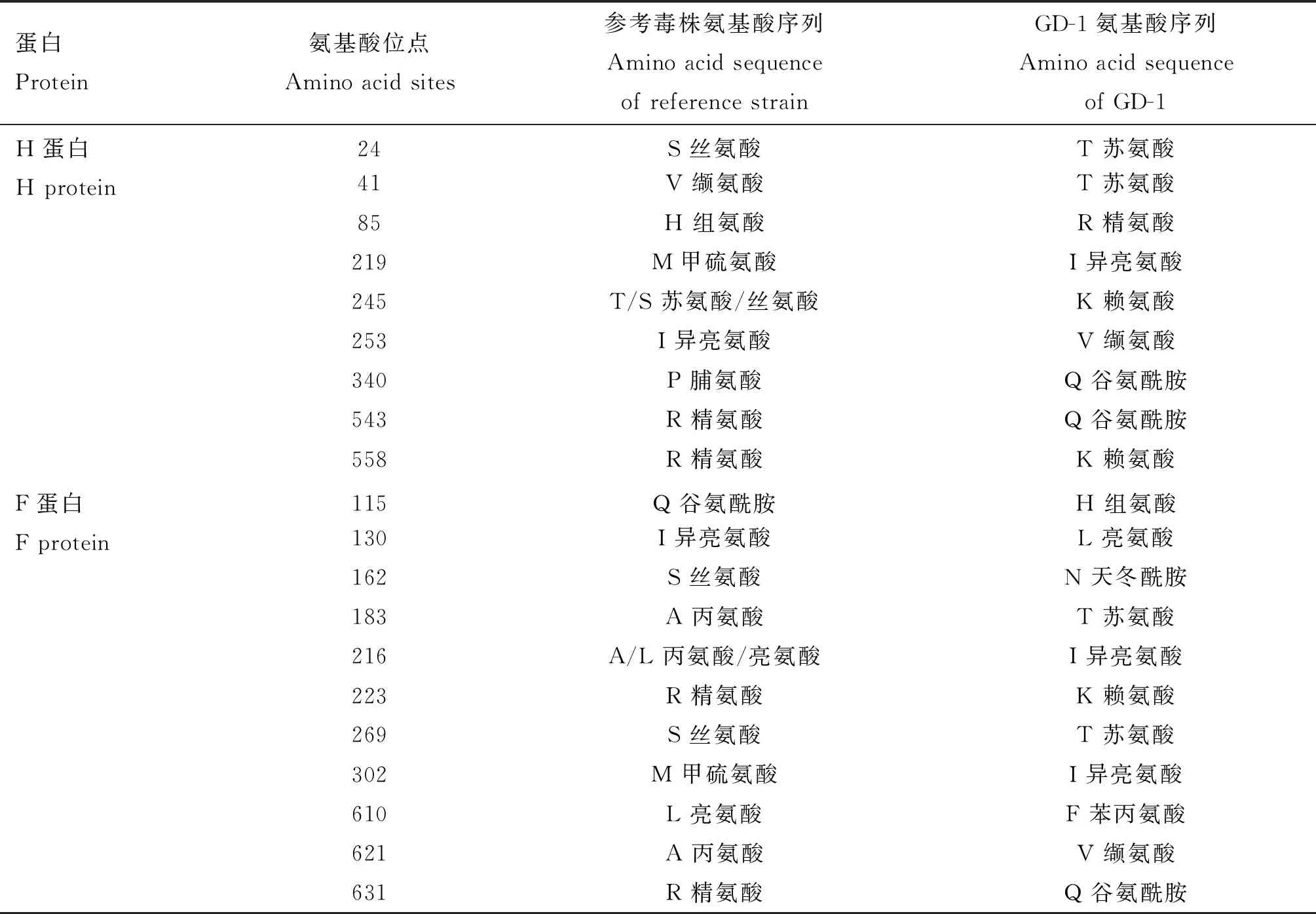
表2 分离株H、F蛋白氨基酸变异位点分析
3 讨 论
CDV作为引起小熊猫大批量死亡、数量骤减的重要病原之一,其导致的CD暴发具有发病急、传播快、发病率高、死亡率高等特点[14-15]。近年来,各地不断有圈养野生动物感染CDV的报道,如1991—1992年北美动物园的虎、豹等,1994年坦桑尼亚塞伦盖蒂国家公园的狮子,1997年中国重庆动物园的大熊猫、2004年俄罗斯博克罗夫卡的西伯利亚虎,中国广西人工饲养场恒河猴等[16-21]。CDV的宿主范围也越来越广,由传统宿主犬科、浣熊科、鼬科动物扩大至食肉目所有8个科的动物,给疾病的预防增加了难度[22-24]。本研究中的小熊猫,此前未接种过相关疫苗,且长期封闭圈养,排除疫苗免疫造成的感染,更大可能性来自于野毒感染,病例发生之前有针对小熊猫分型研究的团队在该地进行过采样,据了解,研究人员此前的采样地是成都某小熊猫圈养地,而此地此前就发生过小熊猫感染CDV的病例,究竟两地之间的病例是否存在关联还在进一步研究中。
本分离株的H蛋白具有8个潜在糖基化位点(19、149、309、391、422、456、587、603),包含野毒株所特有的309—311糖基化位点,未发现新增的糖基化位点,这与大量文献中报道的野毒株含有8或9个糖基化位点相一致,与当前流行的Asia-4型的CDV一致。根据H、F抗原表位分析,GD-1株的抗原表位与疫苗株的相应序列存在较大差异,蛋白抗原决定簇发生改变,暗示更多免疫逃避株的存在。与各参考毒株相比,该株H、F蛋白还存在不同数量的氨基酸位点变异,这些关键位点氨基酸的变异,是否影响了病毒本身传播能力或毒力还有待进一步研究。
基于犬瘟热病毒的地域流行情况,目前已报道过约17个基因型[25-29],分别称为亚洲1~4型、美国1型(多为疫苗株)、美国2~5型、欧洲1型(南美洲1型)、欧洲野生动物型、南美洲2型、南美洲3型、北极型、类洛克伯恩型、非洲1型、非洲2型等。构建基于F、H基因进化树,GD-1株在基因型判定为Asia-4,但是与Asia-4的毒株基因序列仍存在差异,隶属小的进化分支。与当前我国流行的Asia-1型各野毒株亲缘关系较远,之前本研究团队报道过广东某地分离小熊猫源CDV,均属于Asia-1型[30-31]。据Piewbang等[26]近年的报道,Asia-4基因型主要在泰国地区呈流行趋势,宿主源主要是犬、果子狸以及新报道的麝香猫等。本研究结果提示在高强度的疫苗免疫之下,国内犬瘟热病毒免疫逃避株越来越多,病毒本身发生更多漂移或漂变,而关于本研究中毒株的传播能力、致病机制等有待进一步研究。
4 结 论
本研究在国内首次报道了位于Asia-4的小熊猫源CDV,并对病毒的H、F 蛋白进行系统分析,在目前缺乏野生动物专用犬瘟热疫苗使用的情况下,了解新流行毒株的分子特点及变异情况、对于圈养野生动物犬瘟热的科学防控具有指导意义。
