猪源组织蛋白酶S抑制O型口蹄疫病毒在PK-15细胞复制
2021-06-30史喜绢刘原子张大俊申超超袁兴国任瑞瑞杜晓华张克山郑海学刘湘涛
史喜绢,刘原子,张大俊,侯 景,申超超,杨 博,张 婷,袁兴国,任瑞瑞,杜晓华*,张克山*,郑海学,刘湘涛
(1.甘肃农业大学动物医学院,兰州 730070; 2.中国农业科学院兰州兽医研究所 家畜疫病病原生物学国家重点实验室 农业部畜禽病毒学重点开放实验室 国家口蹄疫参考实验室,兰州 730046)
口蹄疫(foot-and-mouth disease,FMD)是由口蹄疫病毒(foot-and-mouth disease virus,FMDV)感染偶蹄动物引起的一种烈性传染病[1-3]。FMDV是微RNA病毒科(Picornaviridae)口蹄疫病毒属(Aphthovirus)的成员,已知有O、A、C、Asia1和SAT1、SAT2、SAT3 7种血清型,成熟的FMDV粒子无囊膜,具有二十面体对称性[4-6]。FMDV全基因组约为8 400 bp,含有一个大的开放阅读框(ORF),ORF编码的多聚蛋白被病毒自身编码的蛋白酶(L、2A、3C)切割成4个结构蛋白和8个非结构蛋白[7-9]。FMDV可利用自身编码的蛋白进化形成抑制或逃避宿主先天性免疫反应,从而促进其自身在宿主体内的存活和复制[10]。FMDV前导蛋白酶(LPro)通过降低IFN-α/β和干扰素刺激因子的早期分泌水平,从而阻止宿主蛋白的合成[11]。FMDV 3A蛋白能抑制病毒触发的IFN-β信号通路,从而逃避宿主免疫反应[12]。同样,宿主也可以识别病原体并激发炎症反应来抑制病毒复制[13]。已经发现有一些宿主蛋白在FMDV复制过程中起重要作用,如DCTN3与FMDV 3A结合负调控FMDV复制[14]。
CTSS是半胱氨酸蛋白酶家族中具有内切肽酶活性的重要成员,由N端16 aa信号肽(SP)、前肽和成熟肽331个氨基酸组成的非活化酶原,其活化需要蛋白酶裂解其N端前肽或由各种因素诱导[15]。CTSS酶活性是其发挥功能的关键,如棕榈酸酯抑制组织蛋白酶诱导内皮细胞侵袭从而抗血管生成,部分是通过抑制CTSL和CTSS活性而起作用的[16]。CTSS的N端有3个凹槽(S1、S2 和S3)与底物的特异性结合相关,这一特性也决定了半胱氨酸蛋白酶抑制剂的特异性;该基因C端有一个与底物结合的位点,即S1’,在酶与主要组织相容性复合体二类分子(MHC-Ⅱ)保守区的特异性结合中起关键作用[17]。CTSS主要在树突状细胞、B细胞和巨噬细胞等抗原呈递细胞中表达[18],其参与细胞外基质、抗血管生成肽和黏附蛋白的降解,促进新生血管形成和肿瘤细胞侵袭转移[19]。CTSS通过调节p38 MAPK和JNK1 途径参与甲基原薯蓣皂苷(methyl protodioscin,MP)诱导的细胞凋亡和自噬[20];通过激活NF-κB和caspase-3 从而诱导肝癌细胞凋亡并增加其化学敏感性[21];也可通过激活CD74调控趋化因子CCL2的表达,进而对肿瘤微环境产生影响[22]。目前对CTSS的研究主要集中在自身免疫性疾病[23]、心血管疾病[24]及肿瘤相关疾病[25],目前尚无有关病原体方面的报道。
初乳在抵抗病原感染中具有重要作用[26]。本团队前期应用iTRAQ技术研究发现母猪初乳中CTSS的含量显著高于常乳,但目前对宿主CTSS在病原感染中的作用研究较少,宿主CTSS在FMDV感染中的作用及其调控机制至今尚不明确。为阐明宿主CTSS在FMDV-O感染过程中发挥的作用,本研究探究了FMDV-O感染和宿主CTSS的相互调控作用,发现宿主CTSS能够抑制FMDV-O在PK-15细胞中复制,而FMDV-O感染增加了宿主CTSS酶活性,进一步研究发现CTSS促进FMDV-O诱导的抗病毒细胞因子产生,明确了猪源CTSS抑制FMDV-O复制的初步原因。本研究结果为更深层次探究猪源CTSS在FMDV-O触发的免疫应答中的作用机制奠定了基础。
1 材料与方法
1.1 材料
口蹄疫病毒毒株FMDV O/MYA98/BY/2010、PK-15细胞和FMDV-O抗体由兰州兽医研究所口蹄疫与新发病流行病学团队保存;兔多克隆抗体CTSS购于Abcam公司;鼠抗Flag单抗、鼠抗Myc单抗、鼠抗β-actin单抗、HRP标记山羊抗鼠IgG二抗和HRP标记山羊抗兔IgG二抗均购于Thermo Scientific公司。
大肠杆菌DH5α感受态、LATaqDNA聚合酶、限制性核酸内切酶BamHⅠ和XhoⅠ、T4 DNA连接酶、RNA抽提试剂Trizol、5×Prime script RT Master Mix、SYBR Permix ExTaqII和蛋白预染Marker均购于宝生物工程大连有限公司; LipofectamineTM2000转染试剂购于Invitrogen公司;Opti-MEM、0.25% EDTA胰酶和胎牛血清(FBS)均购于Gibco公司;MEM细胞培养液和PBS溶液购于建顺公司;ECL显色剂购于Thermo Scientific公司;NP-40裂解液和PMSF购于碧云天公司;织蛋白酶S试剂盒(货号ab65306)购于艾博抗(上海)贸易有限公司;CTSS干扰序列由上海吉玛制药有限公司合成。
1.2 方法
1.2.1 CTSS真核表达质粒的构建 根据GenBank公布的CTSS基因序列(XM_021089893.1)设计合成CTSS引物,引入酶切位点BamHⅠ和XhoⅠ,以pcDNA3.1为载体,构建pcDNA3.1-CTSS-Myc真核表达质粒,进行PCR扩增、酶切和序列测定。
1.2.2CTSS基因 RNAi序列设计与合成 根据GenBank公布的CTSS基因序列(XM_021089893.1)设计并合成CTSS RNAi序列。分别设计了3对针对CTSS基因的RNAi序列。
1.2.3 细胞瞬时转染和病毒感染 将细胞消化后接种于细胞板中,待细胞长至70%~90%时,将质粒与Lip2000试剂(DNA∶Lip2000=1 μg∶2 μL) 分别加至Opti-MEM中,混合后静置15 min,将Opti-MEM混合物直接加至细胞中,将细胞放于培养箱中培养数小时。用无血清的MEM清洗细胞,用无血清的MEM将FMDV-O稀释至MOI为1.0时感染PK-15细胞,置于37 ℃、5%CO2培养箱孵育1 h之后,弃去病毒液,用含2% FBS的MEM维持液继续培养。在感染后12 h收取2份细胞样品,一份用于RT-qPCR,分别检测CTSS和FMDV-O转录水平的变化,并以猪源GAPDH作为内参;一份用于Western blot,分别检测CTSS和FMDV-O蛋白水平的变化,并以β-actin作为内参。
1.2.4 RT-qPCR 收集细胞样品,采用Triozl法提取细胞总RNA,利用合成好的引物进行绝对定量和相对定量检测[1]。相关定量扩增引物信息见表2。

表2 引物序列信息
1.2.5 CTSS酶活性测定 利用组织蛋白酶S活性检测试剂盒(货号ab65306;Abcam)提供的裂解液裂解细胞,离心取50 μL上清于96孔板,加等量反应缓冲液和10 mmol·L-1Ac-VVR-AFC(CTSS底物),根据试剂盒说明书进行处理。使用SpectraMax M5荧光计在400 nm激发波长和505 nm发射波长下测量荧光。
1.2.6 Western blot 收样并处理细胞样品,加入适量的NP-40裂解液(PMSF 1 mol·L-1);充分裂解后,12 000 r·min-1离心10 min,取上清加入含β-巯基乙醇的5×SDS Loading Buffer,100 ℃变性10~15 min,按20 μL的上样量进行SDS-PAGE凝胶电泳[1],最后用高分辨图像采集系统进行ECL显影并保存结果。
1.2.7 数据分析 所有试验至少重复3次,应用 GraphPad Prism 7软件进行分析并作图,运用独立样品T检验进行统计学分析,*.P<0.05表示数据具有统计学意义,**.P<0.01表示数据具有显著性差异,***.P<0.001表示数据间具有极显著性差异。
2 结 果
2.1 FMDV-O感染PK-15细胞促进内源性CTSS表达
将PK-15细胞铺于35 mm小皿中,待细胞长至80%~90%,用MOI为1的FMDV-O感染PK-15细胞,在0、4、8、12 h后收取细胞样品,利用RT-qPCR和Western blot方法检测内源性CTSS的变化。结果表明FMDV-O感染PK-15细胞后内源性的CTSS蛋白水平(图1A)和转录水平(图1B)均高于对照组,提示FMDV-O感染可促进宿主细胞内源性CTSS的表达。
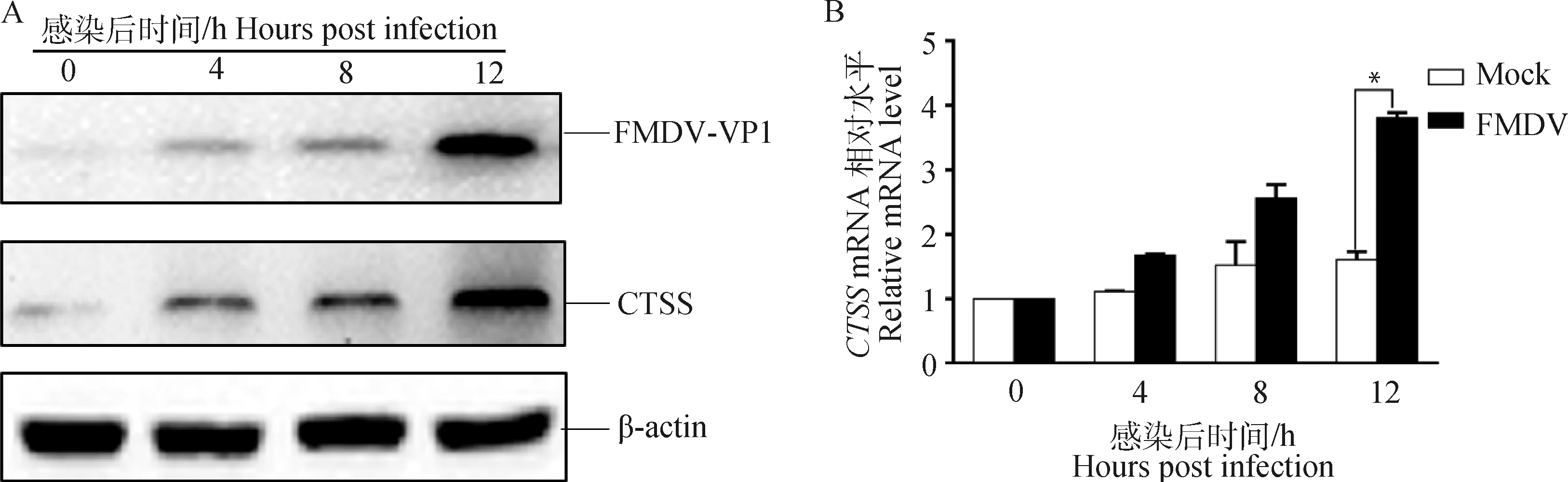
A. Western blot检测CTSS蛋白水平的变化;B. RT-qPCR检测CTSS转录水平的变化;*.P<0.05
2.2 CTSS重组质粒构建及表达验证
构建pcDNA3.1-CTSS-Myc重组质粒,进行PCR扩增,用BamH Ⅰ和XhoⅠ双酶切鉴定,10 g·L-1琼脂糖凝胶电泳检测可在1 000 bp处见目的条带,在5 000 bp 见载体条带(图2A)。经测序后进一步确定该真核质粒构建成功。将构建的pcDNA3.1-CTSS-Myc以不同的剂量分别瞬时转染PK-15细胞,24 h后收取细胞样品,处理样品并进行Western blot验证其表达情况,结果表明pcDNA3.1-CTSS-Myc重组质粒在PK-15细胞中呈剂量依赖性表达(图2B)。

A. 双酶切鉴定结果(M. DL5000 DNA相对分子质量标准;1. BamHⅠ和XhoⅠ酶切产物);B. Western blot验证CTSS蛋白在PK-15细胞表达
2.3 FMDV-O感染能上调CTSS酶活性
转染0.25 μg CTSS重组质粒至PK-15细胞,20 h后用FMDV-O(MOI=1)感染细胞,同时设不用病毒刺激的Mock组,收取0、2、4、6、8、10、12、14 h 细胞样品,裂解细胞用Fluorometric Method检测CTSS酶活性。结果表明,FMDV-O感染能上调CTSS酶活性,且随着FMDV-O感染时间的增加,CTSS的活性也随之增加(图3)。

图3 FMDV-O感染上调CTSS酶活性
2.4 过表达CTSS能抑制FMDV-O复制
转染1、2、4 μg pcDNA3.1-CTSS-Myc至PK-15细胞,转染24 h后用MOI为1的FMDV-O感染细胞,12 h后收取细胞样品,检测其对FMDV-O复制的调控作用,发现随着CTSS表达量增加(图4A、C),FMDV-O复制水平呈现剂量依赖性降低(图4B、C)。结果表明,过表达CTSS抑制FMDV-O在PK-15细胞中复制。

A. RT-qPCR检测CTSS转录水平的变化;B. RT-qPCR检测FMDV-O拷贝数的变化;C. Western blot检测CTSS和FMDV-O蛋白水平的变化; *.P<0.05; **.P<0.01; ***.P<0.001
2.5 CTSS siRNA干扰序列的筛选及其对FMDV-O复制的促进作用
为进一步确定宿主CTSS对FMDV-O复制的影响,针对CTSS设计合成3对特异性siRNA(表1),将siRNA-2947、siRNA-3629、siRNA-3458分别转染至PK-15细胞, 在24和36 h分别收取样品,用RT-qPCR方法选择干扰效果最好的序列,结果显示编号siRNA-2947的干扰效果最好(图5A)。在此试验结果基础上,PK-15细胞转染siRNA-2947,以NC siRNA为对照,36 h后用等量FMDV-O(MOI=1)感染细胞,12 h后收取细胞样品,用RT-qPCR和Western blot检测siRNA-2947对FMDV复制的影响,结果表明siRNA-2947能下调宿主CTSS的表达进而促进FMDV-O在PK-15细胞中复制(图5B、C)。

表1 干扰序列信息

A. RT-qPCR检测CTSS转录水平的变化;B. RT-qPCR检测FMDV-O拷贝数的变化;C. Western blot检测CTSS和FMDV-O蛋白水平的变化; *.P<0.05; **.P<0.01; ***.P<0.001
2.6 过表达CTSS促进FMDV-O诱导的宿主抗病毒细胞因子产生
为明确猪源CTSS抑制FMDV-O复制的原因,检测宿主CTSS是否影响由FMDV-O感染诱导的抗病毒细胞因子产生。在PK-15细胞中分别转染CTSS和pcDNA3.1,24 h后感染FMDV-O(MOI=1),并设无FMDV-O感染对照组,12 h后收集细胞处理样品。RT-qPCR结果显示,CTSS可促进FMDV诱导的IFN-α、IFN-β、IFN-γ、IL-6、IL-10、IL-4的mRNA水平,说明宿主CTSS能激活FMDV-O诱导的宿主抗病毒细胞因子产生。
*.P<0.05; **.P<0.01; ***.P<0.001
3 讨 论
CTSS是一种溶酶体蛋白酶,主要在抗原呈递细胞中表达,其活性调节对于MHC-Ⅱ信号传导及CD4+T细胞介导的免疫反应激活非常重要[27]。有研究报道CTSS活性可以由肠道菌群调节,共生体触发生理性CTSS活性;病原体引起病理性CTSS活性增加,导致T细胞活化和增殖[28]。而本研究发现FMDV-O感染PK-15细胞可上调内源性CTSS的表达并增强CTSS活性。
FMDV以其自身优势拮抗宿主免疫应答进而达到成功感染宿主的目的,当然其生命周期也受不同宿主因素影响[29-32]。有文献报道热休克蛋白DNAJA3与VP1互作并通过自噬/溶酶体途径降解VP1,从而减弱VP1对IFN-β信号通路的拮抗作用,最终抑制FMDV复制[30]。本研究发现FMDV-O感染PK-15细胞可上调内源性CTSS的表达并增强CTSS活性,过表达CTSS抑制FMDV-O在PK-15细胞中复制,而下调内源性CTSS能促进FMDV-O复制。有报道称CTSS能使CX3CL1与CX3CR1相互作用,将免疫细胞募集到炎症部位增加CX3CL1脱落进入间质,从而改变自身免疫性泪腺炎和泪腺分泌[33];干燥综合征患者泪液中CTSS活性升高可诱导促炎细胞因子产生[34];缺乏CTSS会增加高血压小鼠线粒体的受损并提高ROS水平和NF-κB活性,从而调节心脏炎症和纤维化[35]。笔者随后检测了CTSS对抗病毒细胞因子mRNA水平的影响,RT-qPCR结果显示CTSS可促进FMDV-O诱导的IL-6、IL-10、IL-4、IFN-α、IFN-β、IFN-γ基因的转录。这一结果与文献报道一致,提示CTSS可能参与炎症反应。天然杀伤细胞(NKT)可募集并激活其他先天免疫细胞,从而调节多种免疫反应,以加剧肝的炎症反应,但CTSB和CTSS抑制剂可降低LPS诱导的炎症过程中NKT细胞的活化[36],进一步说明CTSS可能与炎症有关。FMDV在进化过程中获得了许多逃避宿主免疫系统的策略[37], 例如FMDV 3A通过破坏RIG-Ⅰ、MDA5和VISA蛋白的表达而抑制病毒触发的IFN-β信号通路[12];FMDV VP3降解JAK1以抑制IFN-γ信号转导途径[38]; LPro可以抑制天然免疫下游抗病毒细胞因子的产生从而促进病毒的复制[39-41],本文虽然检测了抗病毒细胞因子的变化,但机体的免疫系统是一个错综复杂的网络,宿主除了通过调控干扰素信号通路发挥抗病毒功能;还可以通过自噬和凋亡途径影响病毒复制,比如PCBP2和FMDV VP0互作可以通过凋亡途径促进FMDV复制[42]。这说明CTSS促进FMDV-O诱导的抗病毒细胞因子的产生,可能是宿主CTSS抑制FMDV-O复制的原因之一,具体详细机制还需要进一步研究。
本研究首次证实了CTSS在FMDV-O感染过程中发挥抗病毒作用的新功能,为宿主 CTSS拮抗FMDV-O感染方面的研究提供了理论依据,也为下一步探究猪源CTSS在FMDV-O触发的免疫应答中的作用积累了素材;此外,本研究结果也提示CTSS可能作为抑制FMDV-O复制的潜在靶点发挥作用。
4 结 论
FMDV-O感染与宿主CTSS之间具有相互调控作用,FMDV-O感染PK-15细胞显著上调内源CTSS表达并增强CTSS酶活性;过表达CTSS能抑制FMDV-O在PK-15细胞中复制,利用特异性siRNA干扰CTSS表达可以促进FMDV-O复制,并且 CTSS能促进FMDV-O诱导的IFN-α、IFN-β、IFN-γ、IL-6、IL-10和IL-4细胞因子上调表达,明确了CTSS抑制FMDV-O复制的初步原因,具体机制将是下一步研究的方向和重点。
