Lignin metabolism regulates lodging resistance of maize hybrids under varying planting density
2021-06-24LIBinGAOFeiRENBaizhaoDONGShutingLIUPengZHAOBinZHANGJiwang
LI Bin,GAO Fei,REN Bai-zhao,DONG Shu-ting,LIU Peng,ZHAO Bin,ZHANG Ji-wang
State Key Laboratory of Crop Biology,Ministry of Science and Technology/College of Agronomy,Shandong Agricultural University,Tai’an 271018,P.R.China
Abstract Hybrids and planting density are the main factors affecting maize lodging resistance. Here,we aimed to elucidate the mechanism of the regulation of maize lodging resistance by comparing two hybrids at various planting densities from the perspective of lignin metabolism. Our results showed that compared to lodging-susceptible hybrid Xundan 20 (XD20),lodging-resistant hybrid Denghai 605 (DH605) showed a lower center of gravity and culm morphological characteristics that contributed to the higher lodging resistance of this hybrid. Lignin content,activities of key lignin synthesis-related enzymes and G-,S-and H-type monomer contents were significantly higher in hybrid DH605 than in hybrid XD20. Stalk mechanical strength,lignin accumulation and enzyme activity decreased significantly with increasing planting density in the two hybrids. While G-type monomers first decreased with increasing planting density but then remained stable,S-type monomers showed a decreasing trend,and H-type monomers showed an increasing trend. Correlation analysis showed that lodging rate was significantly correlated with plant traits and lignin metabolism. Therefore,maize hybrids characterized by high lignin accumulation,high lignin synthesis-related activities,high S-type monomer content,low center of gravity,high stem puncture strength,high cortical thickness,and small vascular bundle area are more resistant to lodging. High planting densities reduce stalk lignin accumulation,relevant enzyme activities and mechanical strength,thereby,ultimately increasing the lodging rate significantly.
Keywords:summer maize,lignin metabolism,stem morphological characteristics,lodging resistance
1.Introduction
Maize is one of the three major staple food crops in China. Consequently,high maize yields and yield stability are of great significance in ensuring food security for the country. In 2015,the Ministry of Agriculture and Rural Affairs of the People’s Republic of China raised the issue of grain structure contradictions and requested an effort to reduce the maize planting area (http://www.moa.gov.cn). Under conditions of reduced planting area,increasing planting density has become an important measure to ensure high and stable maize yields in China. However,excessively high planting densities will lead to poor light transmission within the crop canopy,which in turn will cause increased plant height and thin,weak stems,thus ultimately leading to increased lodging and yield losses (Tianet al.2016;Tanget al.2007).Furthermore,the cropping season in the Huang-Huai-Hai region makes maize plants prone to extensive lodging due to flooding caused by downpours brought by typhoons. Lodging may cause severe damage to the canopy and photosynthesis may decrease significantly,whereby,the accumulation of photosynthetic products may be strongly reduced and the transport of photosynthetic products severely hindered. Depending on the extent of damage,plants may die,but even if they survive,mechanized harvesting becomes impossible and harvesting costs are greatly increased (Huanget al.2015). Therefore,improving resistance to lodging is an important prerequisite for ensuring high maize yields and yield stability in this region.
Previous studies have shown that under conditions of high planting density,stalk damage due to lodging mainly occurs in the basal three to four internodes. Morphological indexes,such as stalk puncture strength,stem diameter and internode length,are reported to correlate significantly with lodging resistance (Chenget al.2011;Penget al.2014). Thus,maize hybrids resistant to lodging are characterized by thicker stalks with higher puncture strength and shorter internodes. Therefore,it is important to study the morphological characteristics of the stem in the third to fourth stages of growth to estimate the resistance of maize plants to lodging (Liet al.2010;Wanget al.2020).
The main organic components of maize stalks are cellulose,lignin,pectin,proteins,and sugars. Among them,lignin is a complex phenolic polymer underlying plant cell-wall hardness and the mechanical strength of the stem. Previous studies have shown that lignin content and composition are closely related to plant resistance to lodging (Zhenget al.2017);similarly,Muhammadet al.(2018) showed that the physical strength and lignin content of maize stalks are closely related to lodging resistance. Furthermore,lignin accumulation is significantly higher in wheat hybrids with strong flexural strength than in hybrids with low flexural strength (Chenet al.2018).
Previous studies have shown that maize lignin is mainly composed of three monomer types:G (guaiacyl woodlignin),S (syringyl lignin) and H (p-hydroxyphenyl lignin) (Yanget al.2018). Therefore,it is important to determine the content of each of these three monomer types in the stalk to determine the relationship between the lignin composition and maize resistance to lodging.
Although the effects of internode characteristics on maize resistance to lodging have been extensively studied,the effects of lignin synthesis and metabolism have been reported only rarely. Thus,this study mainly focused on using lignin metabolism to explore how hybrid and plant population density might regulate maize resistance to lodging in order to provide breeders with novel insights to improve the efficiency of breeding efforts for resistance to lodging,and to provide theoretical support for field management of maize hybrids that are resistant to lodging.
2.Materials and methods
2.1.Materials and experimental design
These experiments were conducted from 2017 to 2018 at the State Key Laboratory of Crop Biology and at the Experimental Farm of Shandong Agricultural University,China (36°16´N,117°16´E). Total rainfall during the crop cycle,daily average temperature and daily mean wind speed were recorded as 484.4 mm,25.65°C and 1.58 m s-1,respectively (Fig.1). The soil at the study site is a brown loam. Prior to the experiment,total nitrogen in the topsoil layer (0-20 cm) was 0.80 g kg-1,organic matter was 12.26 g kg-1,available phosphorus,available potassium and alkali N were 44.12,85.22 and 116.51 mg kg-1,respectively.
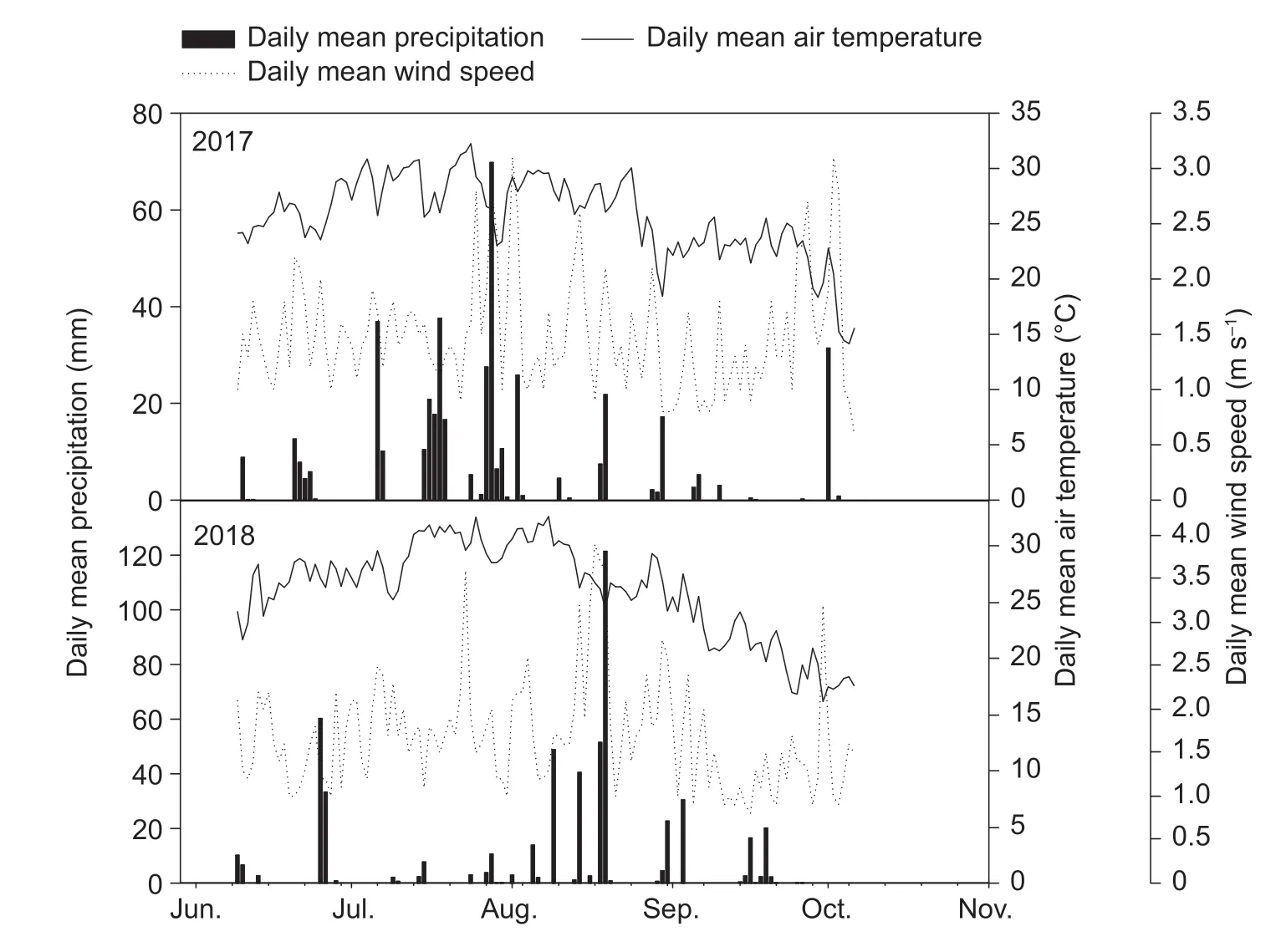
Fig.1 Daily mean air temperature,precipitation and wind speed during the growth periods of 2017 and 2018.
Maize hybrids Xundan 20 (XD20,lodging susceptible) and Denghai 605 (DH605,lodging resistant) were used as experimental materials. Planting density treatments were 60 000,75 000 and 90 000 plants ha-1. The experiment was laid out in a randomized complete block design with three replicates. Plots were 3 m×10 m (30 m2) in size. A row spacing of 60 cm was used for all planting densities,while plant spacings within rows were set at 27.8,22.2 and 18.5 cm,for 60 000,75 000 and 90 000 plants ha-1,respectively.
A total of 300 kg ha-1N (urea,46% N),120 kg ha-1P (superphosphate,17% P2O5) and 240 kg ha-1K (potassium chloride,60% K2O) were applied aiming at a yield of 12 000 kg ha-1. Nitrogen fertilizer was split into two applications,40% at the jointing stage and 60% at the 12th leaf stage. Total P and K doses were applied in a single application at the jointing stage.
2.2.Sampling and measurements
Lodging rateThe ratio of the number of stems displaced from proper and vertical placement in each plot (<30° to the ground) to the total number of plants in the plot was used as lodging rate under natural conditions. Additionally,plant growth stage at the time of lodging occurrence was recorded.
Plant height and center of gravityTen uniform representative plants were selected in each plot at the milk stage (R3). The gravity center of each plant was identified by placing the maize stalk on an overhanging fulcrum and moving the stalk along the fulcrum until the equilibrium point was reached,and that point of the stalk was considered as its center of gravity. The height of erect growth was considered as plant height.
Stalk rind penetration strengthTen uniform representative plants were selected in each plot at R3 and stem puncture strength of the third internode above the ground was determined by a YYD-1 stem strength tester (Zhejiang Top Instrument Co.,Ltd.,Hangzhou,China).
Stem length and diameterTen uniform representative plants were selected in each plot at R3 and the length of the third internode above the ground was measured with a measuring tape. The stem diameter (wide side) at the middle point of the third internode above the ground was measured with a Vernier caliper.
Stem microstructureFive main stems were selected in each plot at R3 and the middle 2 cm portion of the third internode above the ground was fixed with Carnot. Handslicing and saffron staining were used to observe and photograph the structure of vascular bundles in the stalk using a fluorescence microscope camera system (Nikon,Japan). Cortex thickness as well as the number of small and large vascular bundles,and the area of small vascular bundles were measured using the measurement function of that camera system.
Determination of lignin contentTen uniform representative plants were selected in each plot at the 12th leaf stage (V12),tasseling stage (VT) and R3,according to the method described by Chenet al.(2011). Middle portions of the third internode from plants with similar growth were ground to a fine powder in a mortar;liquid nitrogen was then added to the powder,and 0.5 g of this was weighed into a 10-mL centrifuge tube. Chlorophyll was extracted with 95% ethanol;n-hexane:ethanol (2:1,v/v) was used to extract lipids. The remaining precipitate was added to 2.5 mL of 25% bromoacetyl glacial acetic acid;the reaction was maintained at 70°C for 30 min. The samples were cooled to room temperature in cold water,0.5 mL of each sample was transferred to a 50-mL centrifuge tube,and 0.9 mL of 2 mol L-1NaOH and 5 mL acetic acid were added. Then,0.1 mL of 7.5 mol L-1hydroxylamine hydrochloride was added to terminate the reaction. Acetic acid was added to adjust the volume up to 15 mL. Absorbance at 280 nm was measured in a spectrophotometer (Shimadzu UV-2450,Japan). Lignin content is expressed as the absorbance at 280 nm mL-1of the solution per unit mass of fresh sample.
Key enzymes of the lignin synthesis pathwayTen uniform representative plants were selected in each plot at V12,VT,R3,and R6 stages of growth. Middle sections (4 cm) from the third stem internode were placed in a freezer at -80°C for determination of enzyme activities.Phenylalanine ammonia-lyase (PAL) activity was determined according to the method of Assiset al.(2001);4-coumarate-CoA ligase (4CL) activity was determined by an enzymelinked immunosorbent assay (4CL ELISA Test Kit,Jianglai,ShangHai);peroxidase (POD) activity was determined according to the method of Moerschbacheret al.(1988).Finally,cinnamyl alcohol dehydrogenase (CAD) activity was determined according to the method of Morrisonet al.(1994).
Lignin monomer contentTen uniform representative plants were selected from each plot at V12,VT,R3,and R6 stages of growth. A total of 4 cm dry middle sections from the third stem internode were ground and passed through a 40 mesh. A 2.0-g sample was weighed and extracted with a NaCl,absolute ethanol,acetone extracting solution to remove water-soluble,fat-soluble,and protein components from the sample;then,a 1:1 (v/v) mixture of chloroform and methanol was added to remove bound lipids from the sample. After extraction,the precipitate was dried to constant weight. Next,3 mL of 2 mol L-1NaOH and 0.5 mL nitrobenzene were added to 0.02 g samples of the precipitate before heating at 170°C for 1 h in a microwave digestion instrument;then to extract the supernatant,4 mL of ethyl acetate was added to 2 700 μL of supernatant,which was mixed and centrifuged at 5 000×g for 2 min.This procedure was repeated once. The supernatant was evaporated to dryness in a rotary evaporator,reconstituted with 6 mL of 5% acetonitrile in water,sonicated for 1 h,and passed through a 0.22-μm organic filter. The samples were then analyzed using a Triple Quadrupole Mass Spectrometer (Waters Corp.,USA) (Zhenget al.2017).
YieldYield was determined according to the method of Gaoet al.(2018). A field survey was used to determine the number of ears per square meter. Thirty ears were then harvested in each plot to determine yield according to the following formula:
Grain yield (kg ha-1)=Harvest ears ha-1×Kernel number per ear×1 000-grain weight (g)/(1-Moisture content%)/(1-14%)
2.3.Statistical analysis
Analysis of variance was performed using SPSS Software 18.0 followed by the least significant difference (LSD) test to assess the differences among treatments at a probability level of 0.05. Pearson correlation coefficient was used to determine the relationship between lodging rate and each indicator with levels of significance of 0.05 and 0.01.
3.Results
3.1.Lodging rate
In 2017,lodging occurred during tasseling stage (VT),while in 2018 it occurred during R3. The lodging rate for DH605 was 10.06% lower than that for XD20 (averaged over all three planting densities tested). In 2018,the lodging rate for DH605 was 39.47% lower than that for XD20 (averaged over all three planting densities tested). The lodging rates of the two maize hybrids increased with increasing planting density.Compared with D6 (Denghai 605 at 60 000 plants ha-1),lodging rates under D4 (Denghai 605 at 75 000 plants ha-1) and D5 (Denghai 605 at 90 000 plants ha-1) decreased by 3.13 and 2.07% in 2017,and by 4.46 and 0.97% in 2018,respectively. However,there was no signifcant difference between D4 and D5. Similarly,compared with X6 (Xundan 20 at 90 000 plants ha-1),the lodging rates under X4 (Xundan 20 at 60 000 plants ha-1) and X5 (Xundan 20 at 75 000 plants ha-1) decreased by 16.10 and 11.16% in 2017,and by 58.06 and 33.00% in 2018,respectively (Table 1).

Table 1 Effects of hybrids and density on summer maize lodging rate
3.2.Center of gravity and plant height
Plant height and center of gravity were significantly affected by planting density and hybrid. Further,the interaction between planting density and hybrid had a significant effect on center of gravity,but did not significantly influence plant height. In 2017,the average height of the center of gravity of the DH605 hybrid was 6.70% lower than that of the XD20 hybrid (averaged over all three planting densities tested). Similarly,in 2018,the average height of the center of gravity of DH605 was 11.73% lower than that of XD20 (averaged over all three planting densities tested) (Fig.2). The plant heights and heights of center of gravity of the two hybrids increased significantly with increasing planting density.Compared with D4,the heights of the center of gravity under D5 and D6 treatments increased by 3.13 and 7.45% in 2017 and by 3.14 and 9.44% in 2018,respectively,while plant heights increased by 2.29 and 3.36% in 2017,and by 2.59 and 3.80% in 2018,respectively. Similarly,compared with X4,the heights of the center of gravity under X5 and X6 increased by 4.04 and 8.08% in 2017,and by 6.00 and 12.01% in 2018,respectively,while plant heights increased by 1.87 and 3.59% in 2017,and by 1.63 and 5.20% in 2018,respectively (Fig.2).
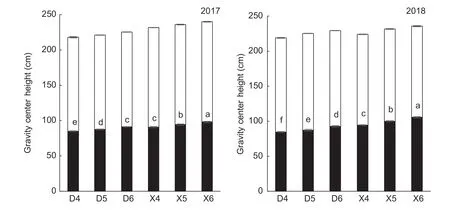
Fig.2 Effects of hybrids and density on plant height and gravity center of summer maize. D4,D5 and D6,Denghai 605 (DH605) at 60 000,75 000 and 90 000 plants ha-1,respectively;X4,X5 and X6,Xudan 20 (XD20) at 60 000,75 000 and 90 000 plants ha-1,respectively. Values are mean±SE (n=5). Different lowercase letters indicate signifcant differences at P≤0.05 level in the same line (LSD test).
3.3.Basal stem morphology and puncture strength
Node length was significantly affected by planting density and hybrid but not by the interaction between the two (Table 2). Planting density significantly affected basal stalk morphology of the maize plants. Thus,compared with DH605,the internode length of XD20 was shortened by 12.04% (averaged over all three planting densities tested) in 2017,and by 16.95% (averaged over all three planting densities tested) in 2018. The internode lengths of the two maize hybrids increased significantly with increasing planting density. Compared with that under D4,mean internode lengths under D5 and D6 increased by 16.49 and 31.95% in 2017,and by 2.63 and 11.40% in 2018,respectively. In contrast,compared with X4,internode lengths under X5 and X6 increased by 22.89 and 42.16% in 2017,and by 16.30 and 17.39% in 2018,respectively (Table 2).
Stem diameter was significantly affected by planting density,hybrid and the interaction between the two (Table 2).Thus,compared with those of XD20,the stems of DH605 were 8.57% coarser in 2017,and 1.29% in 2018 (averaged over all three planting densities tested). The stem diameters of the two hybrids were significantly reduced with increasing planting density. Thus,compared with D4,stem diameters under D5 and D6 were reduced by 14.76 and 21.01% in 2017,and by 14.17 and 22.44% in 2018,respectively.Similarly,compared with X4,the stem lengths under X5 and X6 were increased by 10.77 and 19.82% in 2017,and by 9.01 and 18.28% in 2018,respectively (Table 2).
Stalk rind penetration strength was significantly affected by density and hybrid,but the interaction between the two was not significant (Table 2). Thus,in 2017,stalk rind penetration strength of XD20 was 9.28% lower than that of DH605 (averaged over all three planting densities tested),while in 2018,stalk rind penetration strength of XD20 was 21.54% lower than that of DH605 (averaged over all three planting densities tested). Furthermore,the stalk rind penetration strengths of the two hybrids decreased significantly with increasing planting density.Thus,compared with D4,stalk rind penetration strengths under D5 and D6 decreased by 12.18 and 25.21% in 2017,and by 11.52 and 17.84% in 2018,respectively. Similarly,compared with X4,stalk rind penetration strengths under X5 and X6 decreased by 14.94 and 25.27% in 2017,and by 10.55 and 26.60% in 2018,respectively (Table 2).
3.4.Stem microstructure
Cortex thickness,number of small and large vascular bundles,total number of vascular bundles,and small vascular bundle area,were all significantly affected by planting density and hybrid. However,the effects of the interaction between them on cortex thickness and number of large vascular bundles were not significant. Conversely,such effects were significant on the number of small vascular bundles,total number of vascular bundles,and small vascular bundle area (Table 3). The cortex thickness and small vascular bundle area of XD20 were smaller than those of DH605 under the same planting density,but the number of vascular bundles in XD20 was higher than that in DH605 under low planting density conditions,and the number of small vascular bundles in XD20 under high planting density conditions was lower than that in DH605 (Table 3). Cortical thickness and small vascular bundle area in DH605 were 4.85 and 12.28% higher than those in XD20,respectively (averaged over all three planting densities tested). Cortex thickness,numbers of vascular bundles,and areas of small vascular bundles significantly decreased in the two hybrids with increasing planting density,but the decreases were greater in XD20 than in DH605. Compared with D4,cortex thickness under D5 and D6 decreased by 6.96 and 11.91%,respectively,the total numbers of vascular bundles decreased by 3.36 and 5.52%,respectively,and the areas of small vascular bundles decreased by 17.76 and 17.16%,respectively. Similarly,compared with X4,cortex thickness under X5 and X6 decreased by 7.04 and 15.52%,respectively,the total numbers of vascular bundles decreased by 13.58 and 17.62%,respectively,and the areas of small vascular bundles decreased by 15.15 and 20.40%,respectively (Table 3;Fig.3).
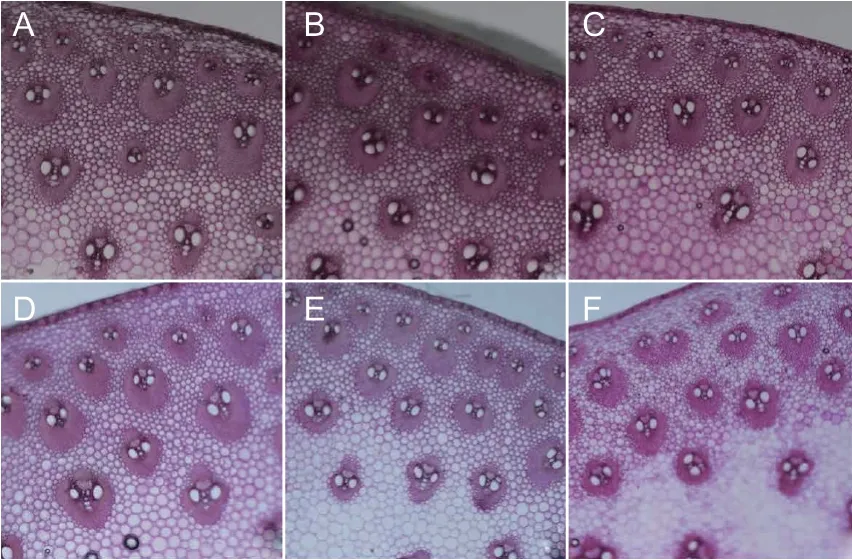
Fig.3 Effects of hybrids and density on the microstructure of summer maize. A-C,D4,D5 and D6;D-F,X4,X5 and X6.D4,D5 and D6,Denghai 605 (DH605) at 60 000,75 000 and 90 000 plants ha-1,respectively;X4,X5 and X6,Xudan 20 (XD20) at 60 000,75 000 and 90 000 plants ha-1,respectively.
3.5.Stalk lignin content
Stalk lignin content was significantly affected by planting density,hybrid and their interaction (Table 4). Lignin accumulation under each treatment gradually increased during growth and peaked at maturity. In 2017,lignin accumulation in XD20 was 9.95% lower than that in DH605 (averaged over all three planting densities tested and growth stages);similarly,in 2018,lignin accumulation in XD20 was 7.33% lower than that in DH605 (averaged over all three planting densities and growth stages) (Fig.4). Lignin accumulation decreased significantly in the stems of the two maize hybrids with increasing planting density,particularly in the stems of XD20. Compared with D4,lignin accumulation under D5 and D6 decreased by 9.75 and 14.28% in 2017 (average of three growth stages),and by 7.02 and 11.64% in 2018 (average of four growth stages),respectively. Similarly,compared with X4,lignin accumulation under X5 and X6 decreased by 13.84 and 22.20% in 2017 (average of three growth stages),and by 6.45 and 13.66% in 2018 (average of four growth stages),respectively.
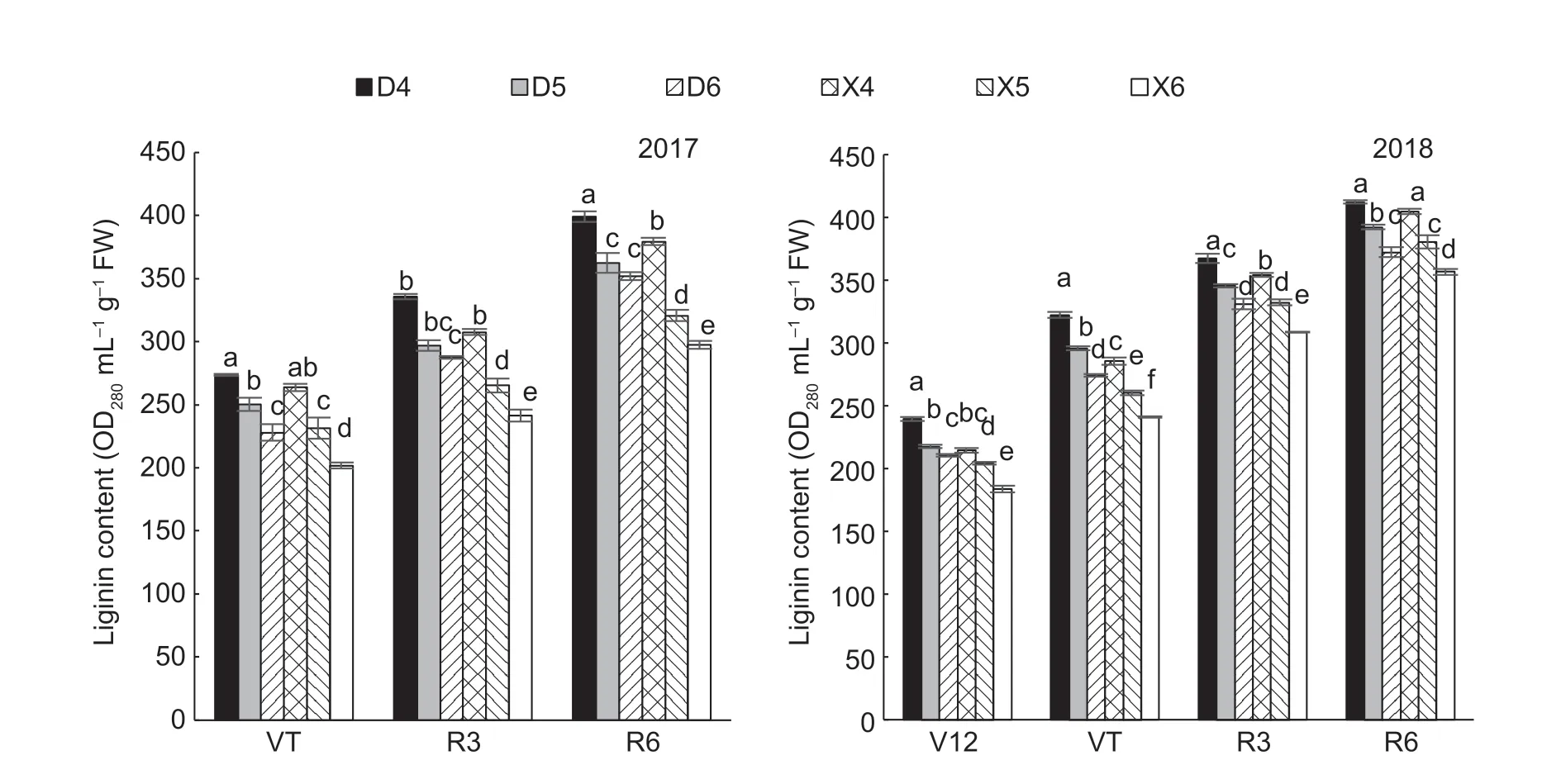
Fig.4 Effect of hybrids and density on lignin content of summer maize. V12,12-leaf period;VT,tassel stage;R3,milk stage;R6,mature period. D4,D5 and D6,Denghai 605 (DH605) at 60 000,75 000 and 90 000 plants ha-1,respectively;X4,X5 and X6,Xudan 20 (XD20) at 60 000,75 000 and 90 000 plants ha-1,respectively. Values are mean±SE (n=3). Different lowercase letters indicate signifcant differences at P≤0.05 level in the same line (LSD test).
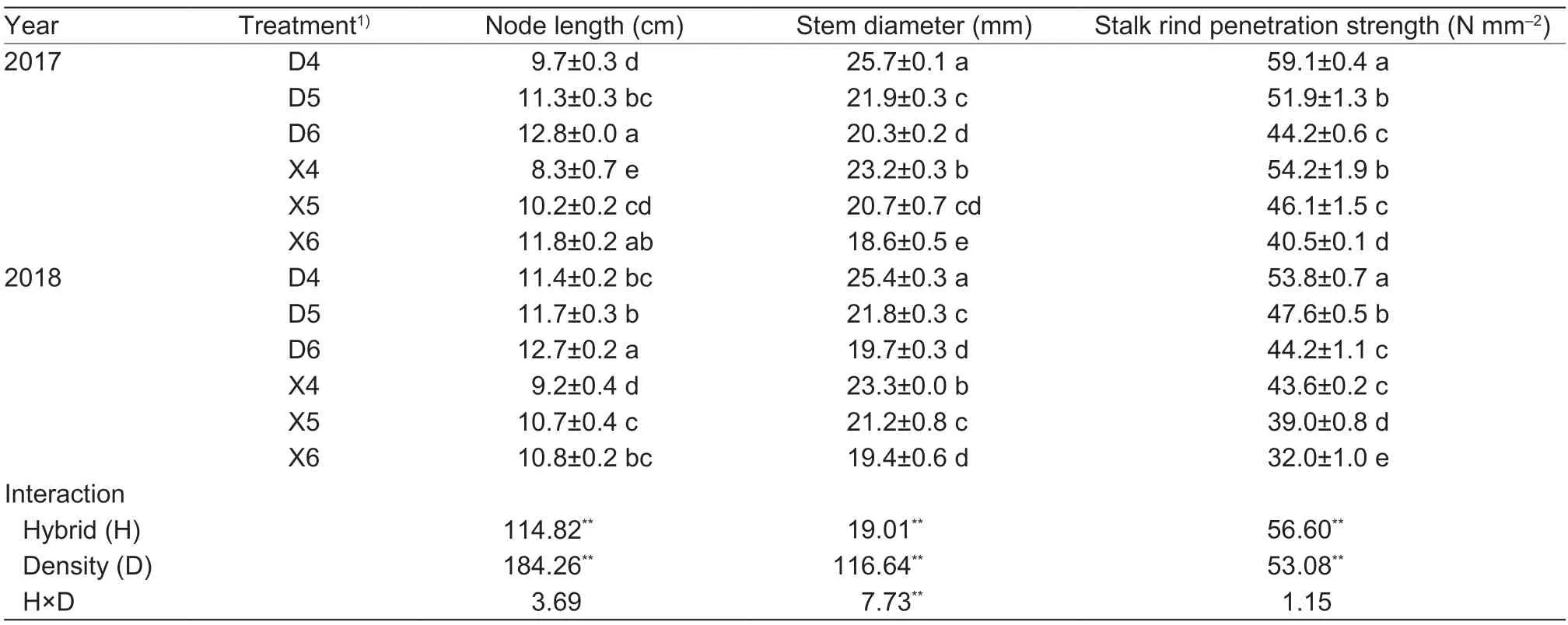
Table 2 Effects of hybrids and density on stem morphology and puncture strength of summer maize base
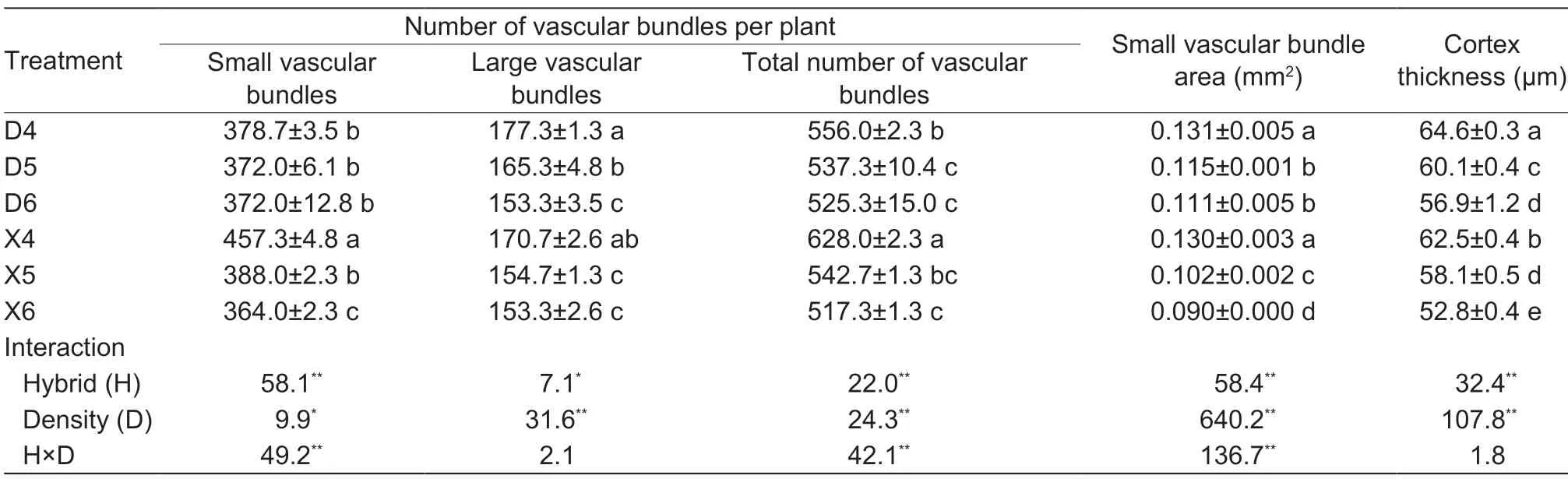
Table 3 Effects of hybrids and density on the microstructure of summer maize
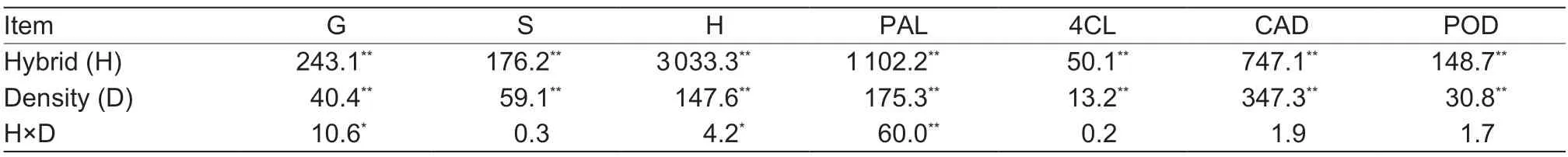
Table 4 Results of ANOVA on the effects of plant density (D) and hybrid (H) on maize lignin monomer components and lignin synthetase activity1)
3.6.Stem lignin-synthetase activity
Planting density and hybrid significantly affected lignin synthesis-related enzyme activity in maize stems. Thus,lignin-synthetase activity was significantly affected by planting density and hybrid,and PAL activity decreased from V12 to R6 in both hybrids. Further,under the same density conditions,PAL activity was higher in DH605 than in XD20 and decreased in both hybrids with increasing planting density,particularly in the latter. Compared with D4,PAL activity under D5 and D6 treatments decreased by 8.54 and 14.40%,respectively (average of four growth stages). In contrast,compared with X4,PAL activity decreased by 14.34 and 35.91% (average of four growth stages) under X5 and X6,respectively (Fig.5).Lastly,the interaction between planting density and hybrid did not show a significant effect on PAL.
In turn,4CL activity peaked at VT and decreased significantly at the end of the growing season. Under the same planting density conditions,the activity of 4CL was higher in DH605 than that in XD20. Further,4CL activity decreased significantly with increasing planting density,with the decrease being significantly lower in DH605 plants than in XD20 plants. Compared with D4,4CL activity under D5 and D6 treatments decreased by 9.32 and 11.17%,respectively (average of four growth stages).Similarly,compared with X4,4CL activity decreased by 10.51 and 14.30% under X5 and X6,respectively (Fig.5).
The activity of POD in both hybrids gradually increased from V12 to VT and then gradually decreased. Under the same planting density conditions,POD activity in plants of hybrid XD20 was 12.35% lower than that in plants of hybrid DH605 (average of each treatment). POD activity decreased with increasing planting density in the two hybrids,with the decrease in XD20 being significantly greater than in DH605.Thus,compared with D4,POD activity under D5 and D6 treatments decreased by 7.94 and 44.70%,respectively (average of four growth stages). In contrast,compared with X4,POD activity under X5 and X6 decreased by 11.65 and 16.97%,respectively (Fig.5).
From a maximum rate reached at R3,CAD activity gradually decreased from R3 to R6. In particular,CAD activity in DH605 increased by 11.49% (average of treatments) compared with XD20 under the same planting density condition. CAD activity decreased significantly with increasing planting density and the decrease was significantly greater in XD20 than in DH605. Compared with X4,CAD activity decreased by 10.51 and 14.30% under X5 and X6,respectively (Fig.5). Similarly,compared with D4,CAD activity decreased by 9.32 and 11.70% under D5 and D6,respectively (average of four growth stages). Finally,the interaction between planting density and hybrid did not significantly affect 4CL,CAD or POD (Table 4).
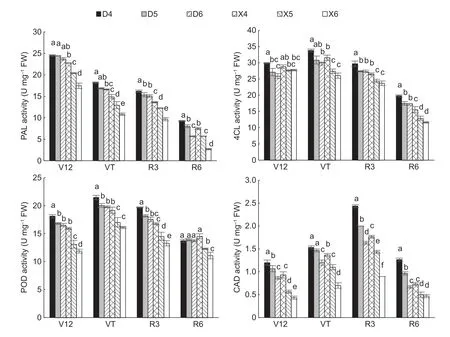
Fig.5 Effects of hybrids and density on lignin synthetase activity in summer maize. PAL,phenylalanine ammonia-lyase;4CL,4-coumarate-CoA ligase;POD,peroxidase;CAD,cinnamyl alcohol dehydrogenase. V12,12-leaf period;VT,tassel stage;R3,milk stage;R6,mature period. D4,D5 and D6,Denghai 605 (DH605) at 60 000,75 000 and 90 000 plants ha-1,respectively;X4,X5 and X6,Xudan 20 (XD20) at 60 000,75 000 and 90 000 plants ha-1,respectively. Values are mean±SE (n=3). Different lowercase letters indicate signifcant differences at P≤0.05 level in the same line (LSD test).
3.7.Stem lignin monomer composition
In maize stalks,lignin is mainly composed of three types of monomers:G,S and H. The proportions of G-and S-monomers are usually larger,whereas the proportion of the H-type monomer is smaller. The accumulation of all three monomers increased as growth proceeded and peaked at maturity.
Stem lignin-monomer composition was significantly affected by planting density and hybrid,but not by the interaction between the two (Table 4). Overall,the accumulation of S-type monomers in the stems of the two experimental hybrids showed a decreasing trend for the same growth stage with increasing planting density.The accumulation of S-type monomers in DH605 was 32.66% higher than in XD20 under the same planting density and,compared with DH605,the accumulation of S-type monomers in XD20 decreased to a greater extent with increasing planting density. Compared with D4,the accumulation of S-type monomers under the D5 and D6 planting density treatments decreased by 7.53 and 15.65%,respectively (average of each growth stage);similarly,compared with X4,the accumulation of S-type monomers under X5 and X6 treatments decreased by 14.65 and 24.60%,respectively (average of each growth stage).
The accumulation of G-type monomers in the stems of both maize hybrids studied here initially decreased and then became stabilized. Under the same planting density condition,the accumulation of G-type monomers in DH605 was 8.38% higher than in XD20. Compared with D4,the accumulation of G-type monomers under D5 and D6 treatments decreased by 9.07 and 9.67%,respectively. Similarly,compared with X4,the accumulation of G-type monomers under X5 and X6 treatments decreased by 7.13 and 10.70%,respectively.
The amounts of accumulated H-type monomers showed different trends at different growth stages. Under the same planting density condition,the accumulation of H-type monomers in DH605 was 69.46% higher than in XD20.During the V12 growth stage,the accumulation of H-type monomers in DH605 decreased with increasing planting density. Compared with D4,the accumulation of H-type monomers under the D5 and D6 treatments decreased by 18.05 and 30.10%,respectively. The accumulation of H-type monomers at VT,R3 and R6 in DH605 increased with increasing planting density. Compared with D4,the accumulation of H-type monomers under D5 and D6 increased by 6.47 and 12.45%,respectively. The accumulation of H-type monomers increased with increasing planting density during the growing season. Compared with X4,the accumulation of H-type monomers under the X5 and X6 planting density treatments decreased by 14.55 and 32.13%,respectively (Fig.6).
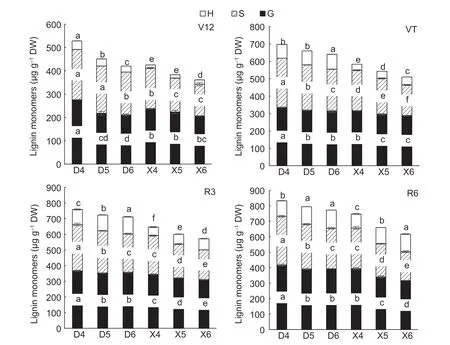
Fig.6 Effects of hybrids and density on lignin monomer components of summer maize (2018). D4,D5 and D6,Denghai 605 (DH605) at 60 000,75 000 and 90 000 plants ha-1,respectively;X4,X5 and X6,Xudan 20 (XD20) at 60 000,75 000 and 90 000 plants ha-1,respectively. H,p-hydroxyphenyl lignin;S,syringyl lignin;G,guaiacyl lignin. V12,12-leaf period;VT,tassel stage;R3,milk stage;R6,mature period. Values are mean±SE (n=3). Different lowercase letters indicate signifcant differences at P≤0.05 level in the same line (LSD test).
3.8.Yield
Number of ears harvested,number of grains per ear,1 000-grain weight and yield were all significantly affected by planting density,hybrid and their interaction (Table 5).The yield of DH605 was 36.02% higher than that of XD20,mainly because the number of harvested ears of DH605 was 29.34% higher than that of XD20. Compared with D4,plant yields under D5 and D6 increased by 6.18 and 12.06%,respectively. Compared with X4,plant yield under X5 increased by 15.66%,but yield under X6 decreased by 37.10%,mainly because the number of ears harvested in X6 decreased by 13.33% compared with X4.
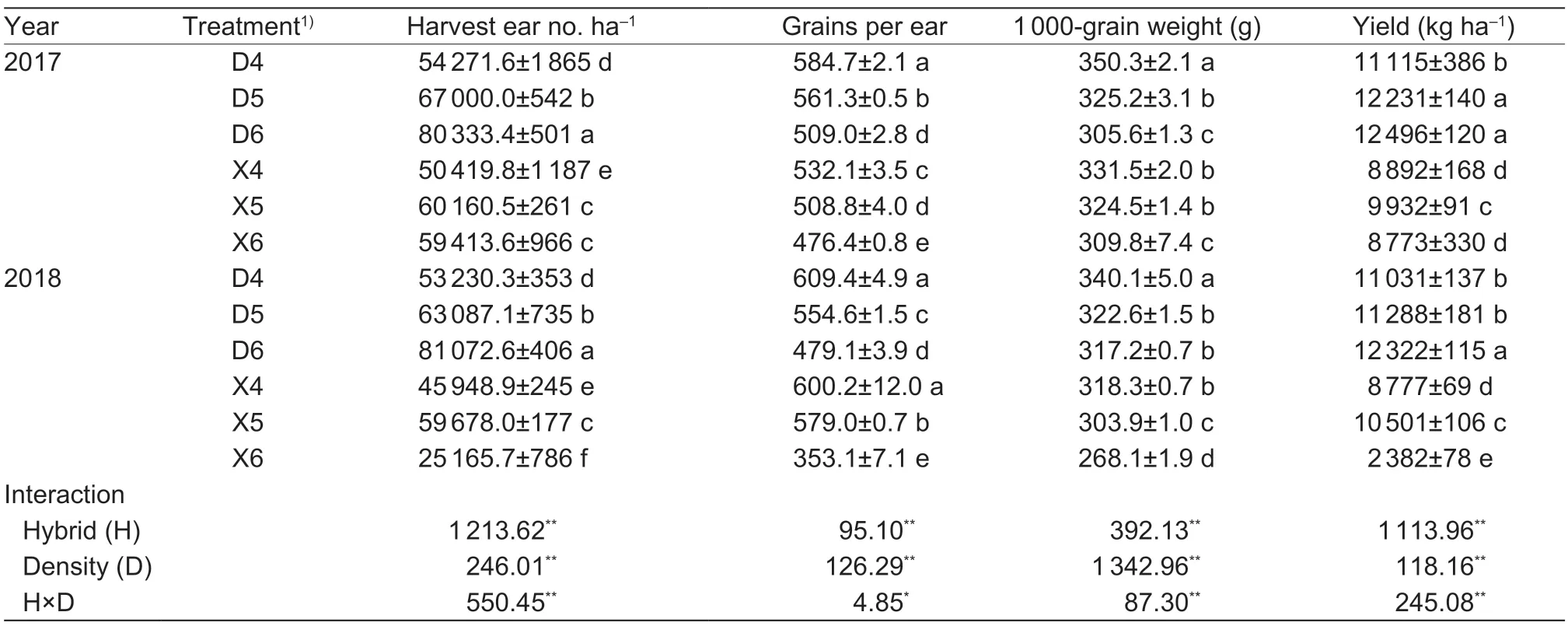
Table 5 Effects of hybrids and density on yield and its components of summer maize in 2017-2018
3.9.Correlation analysis
There was a significant negative correlation between maize lodging rate and lignin content (r=-0860). Furthermore,lodging rate was consistently negatively and significantly correlated with lignin synthesis-related enzyme activities (r-values in the range between -0.878 and -0.965). Lignin accumulation significantly reduced lodging rate. The correlation coefficients for G-and S-type monomers for lodging rate were -0.948 and -0.898,respectively. Although lower,there was also a significant negative correlation between H-type monomer accumulation and lodging rate,(r=-0.570). These observations clearly indicate that lignin metabolism is closely related to lodging resistance in maize.
Significant negative correlations were recorded between maize lodging rate and stem puncture strength,stem diameter,small vascular bundle area,and cortical thickness (r-values in the range between -0.532 and -0.801).Conversely,there was a significant positive correlation between lodging rate and height of the center of gravity,(r=0.933). Coincidently,internode length was significantly and positively correlated with lodging rate (r=0.539). On the other hand,no significant correlation was observed between the number of vascular bundles and lodging rate. Finally,lodging rate was significantly and negatively correlated with yield (r=-0.827) (Fig.7).

Fig.7 Correlation analysis. LP,lodging percentage;PAL,phenylalanine ammonia-lyase;CAD,cinnamyl alcohol dehydrogenase;4CL,4-coumarate-CoA ligase;POD,peroxidase;NL,node length;SD,stem diameter;G,guaiacyl lignin;S,syringyl lignin;H,p-hydroxyphenyl lignin;CG,gravity center;SVBA,small vascular bundle area;CT,cortex thickness;NVB,number of vascular bundles;SPS,stalk rind penetration strength;LC,lignin content. ** and *,significant difference at the 0.01 and 0.05 probability levels,respectively. n=12.
4.Discussion
4.1.Plant stem characteristics and yield
Increasing planting density is an effective method to increase maize yield;however,extremely high planting densities tend to cause maize plants to have a higher center of gravity and a thinner stem,both of which greatly increase the risk of lodging (Wanget al.2012;Okunoet al.2014;Luet al.2015).Previous studies have shown that lodging rate is significantly correlated with the height of the center of gravity (Maet al.2014). In this study,the height of the center of gravity in plants of hybrid DH605 was significantly lower than that of plants of hybrid XD20 at a given planting density,although the center of gravity shifted upwards along the stem in both hybrids with increasing planting density. Correlation analysis showed that the height of the center of gravity of the maize was significantly and negatively correlated with lodging rate.This finding is consistent with previous reports,suggesting that the selection of maize plants with a lower center of gravity is beneficial for maintaining a low rate of lodging at high planting densities.
Previous studies have shown that the physical properties of the stem correlate significantly with maize resistance to lodging. Stem thickness at the basal internodes is significantly and negatively correlated with lodging rate,as reflected by stem puncture strength,an important mechanical index of stalk strength that significantly and negatively correlated with field lodging rate (Gouet al.2010;Liet al.2010;Guoet al.2016;Xueet al.2016).Previous studies have shown that stalk puncture strength was significantly reduced with increasing planting density (Gouet al.2007;Guoet al.2016). In the experiments reported here,stem diameter and stem puncture strength were significantly higher in DH605 than in XD20 at the same planting density. Concomitantly,stem diameter and stem puncture strength of both maize hybrids decreased significantly with increasing planting density. Further,the resulting decrease in mechanical strength was greater for XD20 than for DH605 stems.
Correlation analysis showed that stem diameter and stem puncture strength were significantly and negatively correlated with lodging rate. Cortical thickness,small vascular bundle area and number of vascular bundles were reportedly significantly and positively correlated with maize resistance to lodging (Renet al.2013;Guoet al.2016).Consistently,this study showed that small vascular bundle area and cortical thickness were significantly and negatively correlated to lodging rate,but no significant correlation was found between vascular bundle number and lodging rate. Small vascular bundle area,and cortical thickness and density were significantly larger in DH605 plants than the corresponding values in plants of hybrid XD20,and the small vascular bundle area and cortical thickness of both hybrids decreased with increasing planting density.These decreases in small vascular-bundle area and cortical thickness in XD20 stems were significantly greater than in DH605 stems. Therefore,DH605 showed significantly higher resistance to lodging than XD20. As the values of morphological traits and the stalk mechanical strength of DH605 were significantly greater than those of XD20,the advantages of DH605 stalks become obvious at high planting density.
4.2.Lignin metabolism characteristics
Previous studies have shown that PAL is the rate-limiting enzyme in the synthesis of lignin. This enzyme activity significantly affects the accumulation of lignin (Hahlbrock and Scheel 1989). The 4CL catalyzes the conversion of cinnamic acid derivatives to the corresponding coenzyme A (CoA) esters;in turn,CAD catalyzes the dehydrogenation reaction of lignin monomers corresponding to different organisms.In the final step in lignin synthesis,POD is involved in the polymerization of lignin monomers,and the activities of these four enzymes are significantly correlated with the accumulation of lignin (Yanget al.2015;Kamranet al.2018;Muhammadet al.2018). The results reported herein were consistent with those of previous studies. The key enzymes of the lignin synthesis pathway (PAL,4CL,CAD,and POD) in maize plants all maintained high activity before R3. Hybrid and planting density had significant effects on lignin synthesis. Further,these key enzyme activities in the lignin biosynthesis pathway were significantly higher in DH605 than in XD20 at the same planting density,while they all decreased significantly in both hybrids with increasing planting density,particularly in plants of hybrid XD20. Correlation analysis showed that the four enzyme activities were significantly and positively correlated with maize resistance to lodging.
This study revealed the effects of two contrasting maize hybrids and three planting densities on lignin accumulation and composition,and their relationships with resistance to lodging. Lignin is one of the main structural components in plant stems,as it provides mechanical support to the plant (Chenet al.2011). Previous studies have shown that greater lignin accumulation in stems significantly increases plant resistance to lodging (Wanget al.2015). Consistently,our results showed that lignin accumulation increased gradually during growth and was significantly correlated with plant resistance to lodging. Furthermore,lignin accumulation in plants of hybrid DH605 was significantly higher than that in plants of hybrid XD20 at the same planting density,and it decreased with increasing planting density. However,the accumulation of lignin in DH605 plants was still significantly higher than in XD20 plants at high planting density.
Previous studies have shown that maize stalk lignin is mainly composed of G-,S-and H-type monomers (Yanget al.2018). S-type monomers mainly provide mechanical support,which is closely related to plant resistance to lodging,whereas G-type monomers are mainly related to the transport of materials (Zhenget al.2017). Our results showed that the trend followed by the changes in the total amount of the three types of lignin monomers was basically consistent with the trend followed by the changes recorded in lignin accumulation. The G-and S-type monomers were significantly and negatively correlated with lodging rate.Further,the three lignin-monomer types were significantly higher in DH605 than in XD20 plants at the same planting density. Compared with the G-type,S-type lignin monomers responded more strongly to planting density,while H-type monomers increased with increasing density.
5.Conclusion
DH605 showed greater resistance to lodging regardless of planting density,and plants of this hybrid were characterized by a center of gravity located closer to ground level on the stem,a larger stem diameter,a greater stem puncture strength,a larger cortical thickness,and a greater characteristic size and number of vascular bundles. Additionally,plants of the same hybrid showed higher lignin accumulation,higher lignin synthesis-related enzyme activities and higher content of S-type lignin monomer,as are typical of maize plants with strong lodging resistance. Concomitantly,this hybrid maintained strong stalks and low lodging rates at high planting density. Altogether,these data clearly indicate that breeders should select for these morphological traits while breeding for maize resistance to lodging.
Acknowledgements
The authors gratefully acknowledge the earmarked fund for China Agriculture Research System of MOF and MARA (CARS-02),the National Key Research and Development Program of China (2017YFD0300304) and the National Natural Science Fundation of China (31671629). We also wish to express our gratitude to the reviewers for their vast knowledge and experience in the subject matter of this paper.
Declaration of competing interest
The authors declare that they have no conflict of interest.
杂志排行

Journal of Integrative Agriculture的其它文章
- Effects of plant density and nitrogen rate on cotton yield and nitrogen use in cotton stubble retaining fields
- Adoption of small-scale irrigation technologies and its impact on land productivity:Evidence from Rwanda
- Comparison of grain yield and quality of different types of japonica rice cultivars in the northern Jiangsu plain,China
- Natural nematicidal active compounds:Recent research progress and outlook
- lmproving grain appearance of erect-panicle japonica rice cultivars by introgression of the null gs9 allele
- Comparative transcriptome analysis of different nitrogen responses in low-nitrogen sensitive and tolerant maize genotypes