冷藏卵形鲳鲹优势腐败菌的分离鉴定及致腐能力分析
2021-06-22于淑池冯紫蓝梁宇晴徐小雄裴志胜
于淑池,杨 毅,2,冯紫蓝,梁宇晴,徐小雄,裴志胜,杨 波
(1.海南热带海洋学院,食品科学与工程学院,海南三亚572022;2.中国海洋大学食品科学与工程学院,山东青岛266003)
卵形鲳鲹(Trachinotus ovatus)又名金鲳,系鲈形目鲳鲹属,广泛分布于广东、广西、海南等东南沿海[21,是我国近年来新增的养殖海洋经济鱼种[2];据2019年首届中国金鲳鱼产业高峰论坛数据显示,卵形鲳鲹的养殖产量已突破15万吨以上,产值近100亿元;卵型鲳鲹肉质细嫩,味道鲜美,无肌间刺,是加工鱼块的优良材料[3];同时其富含水分、蛋白质和不饱和脂肪酸,在加工和贮藏过程中易腐败变质,微生物是导致其腐败的主因[4]。
水产品的腐败过程中只有部分优势细菌能大量增殖,在贮藏末期占据主导地位并最终导致产品腐败变质,这些产生腐败代谢产物(臭味、异味、颜色)的优势微生物被称为优势腐败菌(dominant spoilage bacteria)或特定腐败菌(Specific Spoilage Organism,SSO)[5]。在冻结温度以上,水产品特定腐败菌(SSOs)的生长繁殖主导着鱼肉腐败变质的程度[6]。不同水产品中的特定腐败菌存在差异,鱼类捕获后到货架期终点,特定腐败菌的组成因鱼种、捕获季节、水域和贮藏条件等不同而异,但在贮藏条件不变的情况下,每种水产品都有其固定的特定腐败菌[7]。目前已经确定了冷藏条件下大黄鱼、大菱鲆、鲤鱼、罗非鱼、南美白对虾、鹰爪虾、太平洋牡蛎、三文鱼和鲳鱼等多种水产品的特定腐败菌[8],卵型鲳鲹优势腐败菌的研究仅年益莹等[9]采用酸化减菌处理,测定了微冻(-3℃)贮藏条件下金鲳鱼块菌落总数、菌相变化,但4℃冷藏条件下卵形鲳鲹优势腐败菌的研究还未见报道。
特定腐败菌与水产品货架期密切相关[5],是水产品品质控制的关键点;因此确定优势腐败菌对靶向控制其冷藏腐败变质具有重要意义。一种水产品通常只有一种或几种特定腐败菌,在贮藏初期,数量和种类并不占优势,但它们的生长繁殖速度快,到贮藏末期在数量与比重上占绝对优势,且腐败活性强[10],因此SSOs的是贮藏末期的数量优势菌,同时又具备较强的产生异味的定性能力(腐败潜在性)和产生腐败代谢物的定量能力(腐败能力)[11]。近年来,国内外部分学者开始利用腐败菌产生的腐败代谢产物作为其腐败能力的定量指标[12]。TVB-N、生物胺、三甲胺等腐败代谢产物产量因子均可作为水产品优势腐败菌致腐败能力定量分析的评价标准[13]。本研究以卵形鲳鲹为原料,对4℃冷藏卵形鲳鲹贮藏期间腐败细菌进行筛选纯化、菌相分析及部分生理生化指标进行测定,结合16S rDNA技术鉴定腐败菌,以腐败物质的产量因子YTVB-N/CFU为评价标准,定量评价贮藏末期的优势腐败菌的致腐能力,最终确定冷藏卵形鲳鲹的优势腐败菌;以期为卵形鲳鲹腐败菌的靶向控制提供理论参考。
1 材料与方法
1.1 材料与仪器
新鲜卵形鲳鲹(体质量600±10 g)购于三亚市胜利路旺毫超市,保持鲜活状态在30 min内运回实验室;平板计数琼脂培养基(Plate count agar,PCA)、假单胞菌选择性培养基(Pseudomonas CFC Selective Agar,CFC)、铁琼脂培养基(Iron Agar,IA)、结晶紫中性红胆盐葡萄糖琼脂培养基(Violet Red Bile Agar,VRBGA)、乳酸菌分离培养基(Lactobacilli MRS Broth,MRS)、胰蛋白胨大豆琼脂培养基(Trypticase Soy Agar,TSA)、细菌普通培养基(Luria-Bertani,LB) 青岛海博生物技术有限公司;NaCl、HCl、无水乙醇、硼酸 均为分析纯,西陇科学股份有限公司;革兰氏染液试剂盒、细菌基因组DNA提取试剂盒 生工生物工程(上海)股份有限公司;氧化镁 分析纯,山东西亚化学工业有限公司;胰蛋白胨、大豆蛋白胨 奥博星生物技术有限责任公司;酵母浸膏 茂捷微生物科技有限公司;甲基红指示剂 福晨化学试剂厂;亚甲基蓝指示剂 天新精细化工开发中心;过氧化氢 分析纯,富宇精细化工有限公司;PCR扩增试剂及引物华大基因;丙三醇 分析纯,国药集团化学试剂有限公司;孔雀石绿 上海麦克林生化科技有限公司;氧化酶试纸 杭州市西湖区奥图实验仪器经营部。
SQ510C型立式压力蒸汽灭菌器 重庆雅马拓科技有限公司;FA2204B型电子天平 上海天美天平仪器有限公司;LRH-250F型生化培养箱 金坛市盛兰仪器制造有限公司;SWC-FD型洁净工作台苏州安泰空气技术有限公司;Kjeltec2300型凯氏定氮仪 丹麦FOSS公司;icount全自动菌落计数仪杭州迅数科技有限公司;Multifuge X1R型高速冷冻离心机 赛默飞世尔科技(上海)有限公司;BCD-290W型冰箱 青岛海尔股份有限公司;BM2000型电子显微镜 南京江南永新光学有限公司;ZQIY70N台式全温振荡培养箱 上海知楚仪器有限公司;Mastercycler6325PCR扩增仪 EPPENDORF中国有限公司;PowerPac Basic电泳仪、Universal Hood II凝胶成像仪 BIO-RAD中国有限公司。
1.2 实验方法
1.2.1 原料处理 鲜活卵形鲳鲹用冰水混合物致死,清水冲去表面粘液,经去头、去尾、去内脏“三去”处理后,取背部肌肉,切成(50±0.2)g鱼片,放入托盘中覆盖保鲜膜,置于(4±0.5)℃冰箱冷藏6 d[14];分别于初期(0 d)、中期(3 d)、后期(6 d)取贮藏的鱼片进行腐败细菌的分离纯化及鉴定。
1.2.2 细菌分离纯化 分别于4℃冷藏的初期(0 d)、中期(3 d)、后期(6 d),取4℃冷藏卵形鲳鲹鱼片5 g左右置于超净台内,在无菌操作环境下放入灭过菌的研钵中,加入海砂将鱼肉研碎,加无菌生理盐水45 mL,摇床振荡30 min后,取l mL进行10倍梯度稀释,选择3个合适的稀释度(10-4、10-5、10-6),每个稀释度取100μL菌液,均匀涂布于6种不同选择培养基上,其中PCA、IA、CFC、TSA等培养基选择30℃培养48 h,MRS、VRBGA培养基37℃培养48 h,挑选典型菌落,分别在各菌相应的培养基上反复进行平板划线分离,至少3次划线分离后得到纯化单菌落,观察菌落特征,并进行革兰氏染色、芽孢染色、接触酶测试、氧化酶测试,具体的实验方法参照《伯杰氏系统细菌学手册》[15]和《常见细菌系统鉴定手册》[16];挑取典型单菌落,于LB液体培养基中30或37℃摇床培养至对数生长期后,菌悬液加20%甘油于-80℃冰箱中冻藏[17],用于进一步进行16S rDNA基因的提取分析。
1.2.3 16S rDNA分析 将贮藏初期(0 d)、中期(3 d)、后期(6 d)分离纯化得到的腐败菌,经过LB液体培养基培养至对数生长期后,4℃、9000 r/min离心1 min,收集菌体,采用DNA提取试剂盒提取分离纯化所得菌株的DNA,进行PCR扩增,通用引物为27f/1492r,参照董文霞等[18]的扩增条件,95℃预变性5 min后持续变性30 s,45℃退火30 s,72℃延伸30 s,共30个循环,最后72℃延伸30 s;扩增后的产物经1%琼脂糖凝胶电泳确认条带后,送往生工生物工程(上海)股份有限公司(海南分公司)测序,结果登陆NCBI网站进行同源性分析,采用MEGA6.0构建系统发育树。
1.2.4 菌相变化分析 在方法1.2.2中,对贮藏初期、中期、后期的腐败细菌进行分离纯化的过程中,同时选取菌落数在30~300(Colony-Forming Units,CFU)的平板,根据菌落形态特征并结合镜检,对相同菌落细菌进行分组和归类计数,并计算同种菌落细菌占当天菌落总数的比例;再结合16S rDNA基因分析鉴定结果,进一步分析3个贮藏时期的菌相组成及比例变化规律。
1.2.5 优势腐败菌致腐能力定量测定
1.2.5.1 优势腐败菌悬液的制备 将贮藏后期分离纯化的2号腐败希瓦氏菌、5号奥奈达希瓦氏菌、16号霍氏肠杆菌,3株优势菌活化后,取典型菌落接种于LB液体培养基中,希瓦氏菌与霍氏肠杆菌分别置于30、37℃摇床培养,至菌悬液浓度达到108CFU/mL,离心12000 r/min、30 min,弃上清,无菌生理盐水稀释至106CFU/mL,备用。
1.2.5.2 优势腐败菌鱼肉的制备与测定 参照唐文静等[19]法制备无菌鱼块。卵形鲳鲹经“三去”处理后清洗干净,用无菌水冲洗,75%乙醇擦拭卵形鲳鲹鱼体,用无菌剪刀剪背脊内部无污染鱼肉,保证鱼块厚度为1 cm左右、45~50 g/块,制备好的无菌鱼块菌落总数应<102CFU/g[20]。在超净工作台中,将无菌卵形鲳鲹鱼块分为3组,每组3个平行,放入制好的3种菌悬液中,浸泡30 s,捞出沥干,装入无菌的自封袋中置于4℃冰箱中贮藏,每隔2 d取出接种的鱼块进行感官评价、菌落总数、TVB-N指标测定,以未接种菌液的处理作为对照组。
1.2.5.3 测定指标 感官评定参照王者等[21]对卵形鲳鲹的感官评价标准,采用10分制评分法,由10名专业人士组成的感官评定小组,分别从样品的色泽、气味、组织形态和弹性结构4个方面综合评分,结果取平均值。当样品的评定综合指标在6分以下时,视为其初期腐败的开始。
参照GB 4789.2-2016[22]进行菌落总数测定;参照GB 5009.228-2016[23]法测定TVB-N值,参照许振伟等[20]的方法进行腐败能力的定量分析,以腐败代谢产物产量因子(YTVB-N/CFU)为评价指标,计算公式如下:
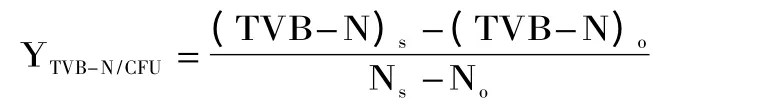
公式中:(TVB-N)0、(TVB-N)s分别表示初始点和货架期终点的TVB-N含量,mg/100 g,N0、Ns分别表示初始点和货架期终点的菌落总数,CFU/g。
1.3 数据处理
采用Microsoft Excel 2010和SPSS 17.0对实验数据进行处理、统计及绘图,数据以平均值±标准差表示;不同优势腐败菌致腐能力的组间显著性分析,采用ANOVA进行分析,不同小写字母表示组间差异显著(P<0.05)。
2 结果与分析
2.1 菌落形态分析
通过6种选择培养基稀释涂布筛选,至少3次以上平板划线分离,卵形鲳鲹4℃冷藏期间共分离出16种单菌落,分别以阿拉伯数字命名,16株腐败细菌菌落形态特征见表1。
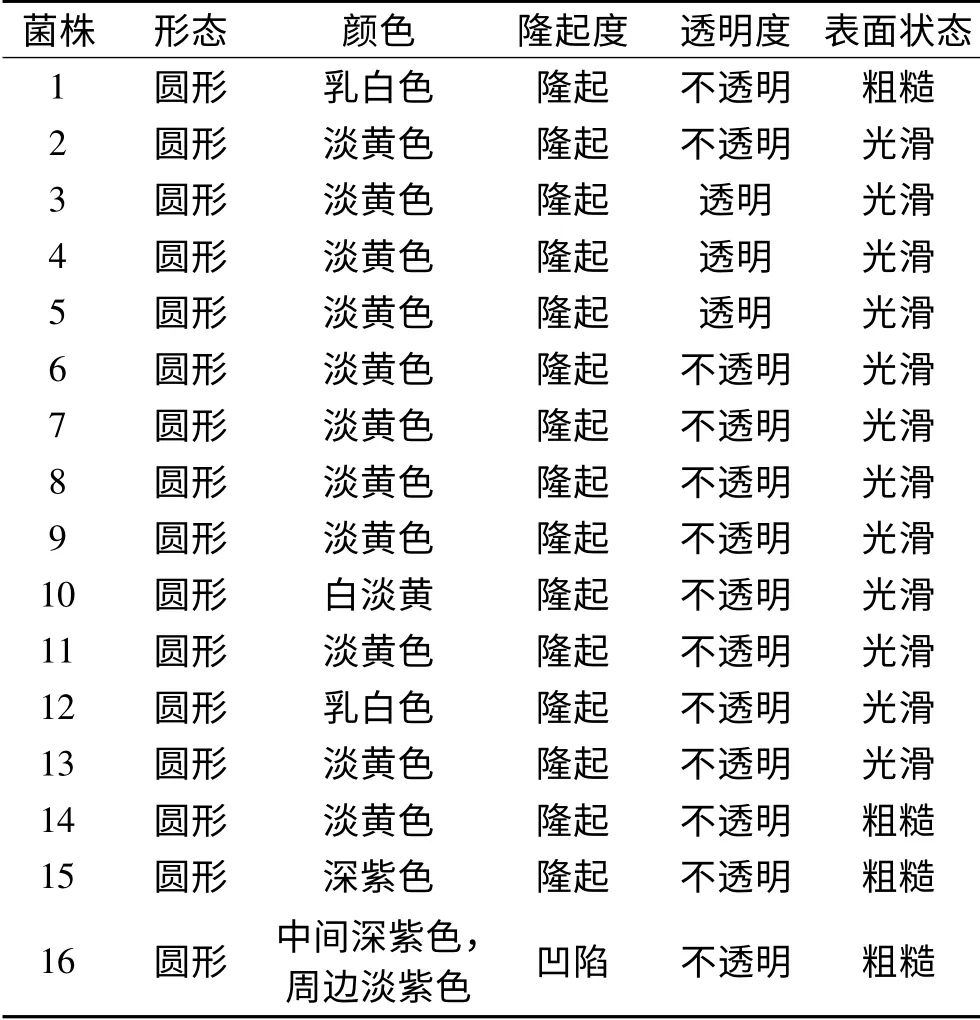
表1 16株腐败细菌的菌落形态特征Table1 Colony morphology of 16 putrefying bacteria
表1可知,16株菌的菌落形态全为圆形;颜色有所不同,大部分为淡黄色,2株为乳白色和2株为深紫色,16号菌株菌落中央深紫色,周边为淡紫色;15株菌菌落表面都为隆起,只有16号菌株菌落凹陷;菌落大部分为不透明,仅有3种菌落透明(3、4、5号);菌落表面状态大多为光滑,只有1和14、15、16号表面粗糙。
2.2 16株腐败细菌形态特征及部分生理生化指标测定结果
表2为分离纯化的16株菌革兰氏染色及其他生理生化特征。
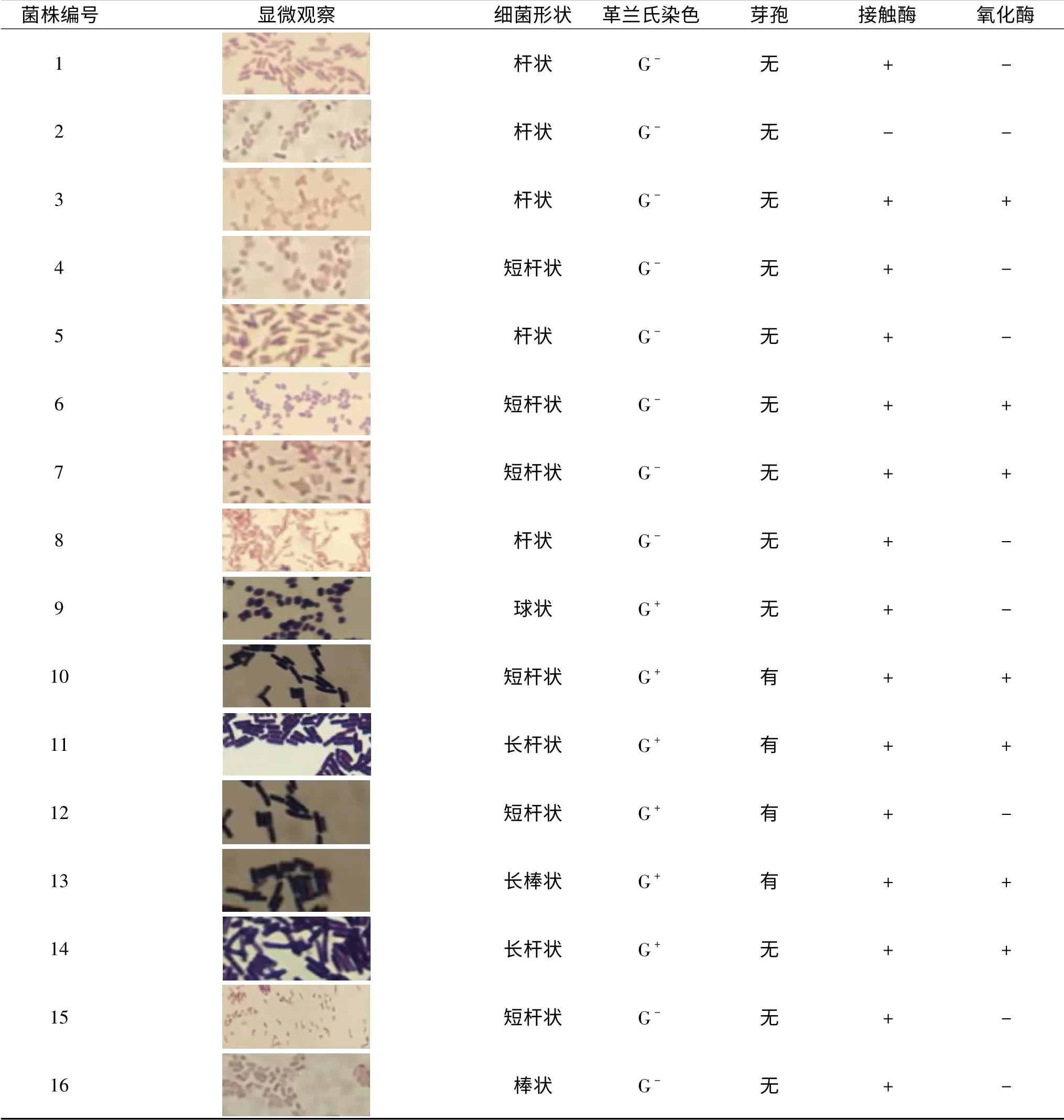
表2 16株腐败细菌的生理生化特征Table 2 Physiological and biochemical characteristics of 16 putrefying bacteria
16株菌中,13株为杆状,1株为球菌,2株为棒状;4株菌(10~13号)有芽孢;此外,除2号菌株以外,其他菌株接触酶实验阳性,说明这些菌株中均存在过氧化氢酶;3、6、7、10、11、13、14号菌株氧化酶实验为阳性,其他均为阴性。革兰氏染色结果表明,16株菌中有6株是革兰氏阳性菌,占37.5%,其余10株为革兰氏阴性菌,占62.5%。
2.3 16S rDNA分析结果
2.3.1 16菌株PCR产物的凝胶电泳检测结果 采用27f/1492r通用引物,对冷藏卵形鲳鲹16株腐败细菌,进行16S全长序列PCR扩增,利用1%的琼脂糖凝胶电泳,检测16S rDNA扩增产物,电泳图谱见图1。
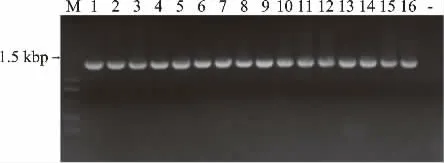
图1 16株腐败细菌的16SrDNA电泳图谱Fig.1 Electrophoretogram of 16SrDNA genes from 16 putrefying bacteria
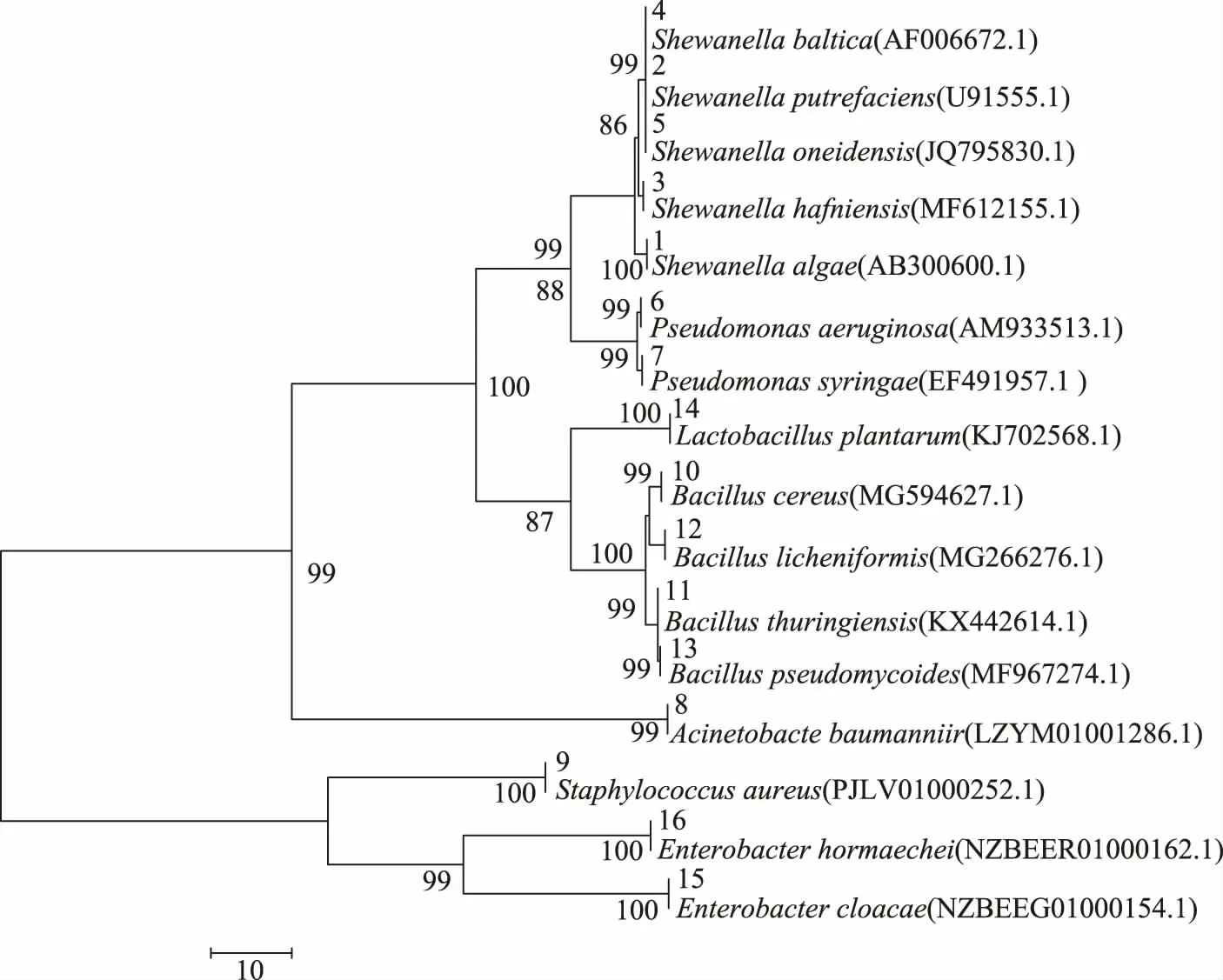
图2 基于16SrDNA序列同源性的16株菌的系统发育树Fig.2 Phylogenetic tree of 16 putrefying bacteria based on 16SrDNA sequences
图1 显示,16株菌PCR扩增产物均得到特异性清晰明亮的条带,片段大小为1.5 kbp左右,显示PCR扩增成功,样品纯度合格,可以用于后期的测序。
2.3.2 16株菌的16S rDNA基因序列分析 将上述PCR扩增产物送测,将检测得到的16株菌的基因序列与NCBI网站中的基因总库进行对比,选取其中同源性(≥98%)最高的菌株序列,用MEGA 6.0构建系统发育树,确定优势腐败菌的种类;结果见图2、表3。
由图2、表3可知,16株菌16SrDNA序列与模式菌株序列的相似度均≥98%,说明鉴定结果可靠。经分子生物学鉴定后,将16株菌的基因序列提交至GenBank获得临时登录号,再与NCBI中的基因总库进行比对,结果16株菌分属于6个菌属,其中5株(菌株编号1、2、3、4、5)属于希瓦氏菌属(Shewanella sp.);2株(6、7号)假单胞菌属(Pseudomonas sp.);8号菌株为不动杆菌属(Acinetobacte baumanniir),9号菌株为葡萄球菌属(Staphylococcus aureus),4株(10、11、12、13号)属于芽孢杆菌属(Bacillus sp.);14号植物乳酸杆菌(Lactobacillus plantarum);2株(15、16号)肠杆菌属(Enterobacter sp.)。

表3 冷藏卵形鲳鲹腐败细菌的16SrDNA序列分析Table 3 16SrDNA sequence analysis of putrefying bacteria in refrigerated Trachinotus ovatus
研究发现[24],鱼类腐败菌的种类受品种、捕获季节、贮藏环境等因素的影响,而有较大的差异,Huang H等[25]发现,鱼类低温冷藏的主要菌相一般为假单胞菌、肠杆菌、芽孢杆菌、乳酸菌、不动杆菌、腐败希瓦氏菌等;一般有氧冷藏动物性产品的腐败细菌主要是蛋白分解能力强的嗜冷性革兰氏阴性菌,包括假 单 胞 菌 属(Pseudomonas sp.)、希 瓦 氏 菌 属(Shewanella sp.)和 肠 杆 菌 科(Enterobacteriaceae)等[26],本研究结果与文献报道相符。
2.4 菌相分析
4℃冷藏卵形鲳鲹贮藏初期(0 d)、中期(3 d)、后期(6 d)的菌相变化见图3。
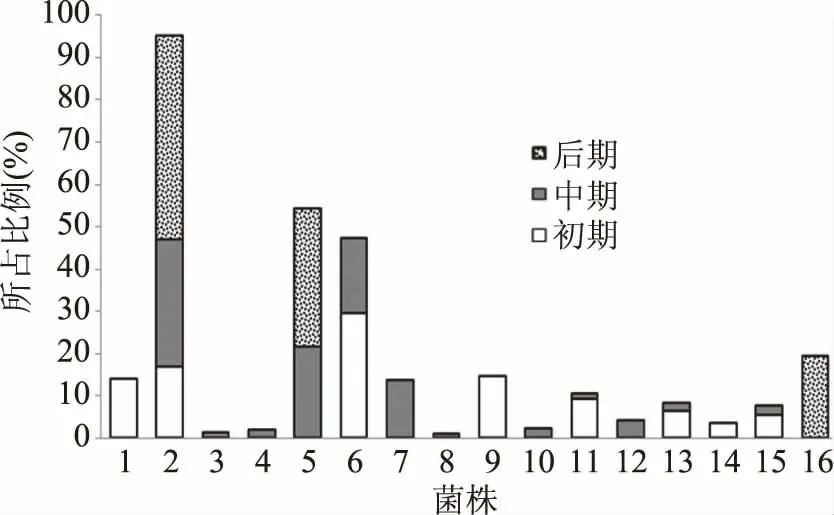
图3 冷藏卵形鲳鲹菌相变化图Fig.3 Phase changes of refrigerated Trachinotus ovatus
不同的选择性培养基选择性分离筛选不同的菌属。IA选择性培养基筛选的是产硫细菌(1~5号菌),主要是希瓦氏菌属;CFC筛选的是假单胞菌属(6~7号菌);PCA培养基为平板计数琼脂培养基(8~9号菌);TSA筛选对营养要求较高的细菌(10~13号菌),主要是芽孢杆菌属;MRS筛选的是乳酸菌属(14号菌);VRBGA筛选的是肠杆菌属(15~16号菌)。
图3可以看出,6种不同的选择性培养基筛选得到的菌株数和比例有所不同,初期(0 d)各菌属均有分布,1号海藻希瓦氏菌(Shewanella algae 14.1%)、2号腐败希瓦氏菌(Shewanella putrefaciens 16.8%)、6号绿脓假单胞菌杆菌(Pseudomonas aeruginosa 29.6%)、9号金黄色葡萄球菌(Staphylococcus aureus 14.8%)占比较高;中期(3 d)时2号腐败希瓦氏菌比例上升到30.4%、5号奥奈达希瓦氏菌比例为21.6%、6号菌株比例下降到17.8%,新出现7号菌株丁香假单胞杆菌(Pseudomonas syringae 13.7%)占比均较高;后期(6 d)筛选到的菌株较少,但比例较高,2号腐败希瓦氏菌比例达到最高48%,5号奥奈达希瓦氏菌26.6%,16号霍氏肠杆菌只在后期出现,占比也较高为19.4%。
从整体上分析,4℃冷藏卵形鲳鲹的菌相构成比较复杂,希瓦氏菌属种类最多、所占比例最高,其次是假单胞菌属。贮藏初期菌相丰富,各菌属均有分布;中期乳酸菌属消失,除希瓦氏菌属和假单胞菌属比例较高以外,其他菌属占比较小;后期,2株希瓦氏菌占比较高(74.6%),其次是肠杆菌(19.4%),其他大部分菌种减少或消失。
年益莹等[9]测定了用HCl酸化减菌处理,对金鲳鱼块微冻(-3℃)贮藏下菌相变化的影响表明,优势腐败菌由贮藏初期的希瓦氏菌过渡到中期的希瓦氏菌、环丝菌及假单胞菌,直到末期的假单胞菌;蓝蔚青等[27]研究发现银鲳冷藏初期细菌多样性较高,优势菌为葡萄球菌(占44.2%)和假单胞菌(占23.9%),而希瓦氏菌只占6.4%,贮藏末期的优势菌则由假单胞菌(占45.7%)和希瓦氏菌(占33.6%)构成。Lalitha[28]研究了养殖罗氏沼虾在冰藏过程中的微生物变化,新鲜的罗氏沼虾中,Enterobacteriaceae和Aeromonadaceae为优势菌;在贮藏末期,40%~52%是H2S产生菌;Dabade等[29]研究了不同冷藏温度下美菲对虾(Penaeus notialis)菌相和优势腐败菌,在28和7℃时,H2S产生菌占优势,而在0℃时,假单胞菌占优势;冷藏卵型鲳鲹的菌相变化与文献报道类似,优势菌也是假单胞菌、希瓦氏菌,只是菌相组成及比例变化不尽相同,贮藏后期希瓦氏菌占绝对优势,说明水产品种类、贮藏条件对菌相组成及比例都有直接影响。
卵形鲳鲹出现了霍氏肠杆菌和芽孢杆菌等,可能是卵形鲳鲹捕获后环境中的腐败菌附着在水产品上所致。Surendran等[30]研究发现热带鱼类肠杆菌含量偏高,郑振霄等[31]也认为冷水鱼冷藏过程中,希瓦氏菌属与肠杆菌属在腐败过程中占主导地位。
卵型鲳鲹贮藏初期革兰氏阴性菌比例为76%,末期上升为97.8%;Lalitha[28]研究了养殖罗氏沼虾在冰藏过程中的微生物变化,新鲜的罗氏沼虾中,革兰氏阴性菌占总菌数的73%,经过19 d的贮藏后,革兰氏阴性菌占总菌数的80%,说明冷藏条件下卵形鲳鲹等水产品的营养基质更适合革兰氏阴性菌的生长。
由菌相分析可知,4℃冷藏卵形鲳鲹贮藏末期希瓦氏菌属(2号腐败希瓦氏菌属、5号奥奈达希瓦氏菌)和16号霍氏肠杆菌3菌株后期所占的比例较大,确定为数量优势菌,进一步进行致腐能力的定量测定。
2.5 优势腐败菌的致腐能力分析
接种3株优势腐败菌后,卵形鲳鲹鱼块冷藏期间,分别测定感官、TVB-N值、菌落总数等指标。
2.5.1 感官评价 接种不同优势腐败菌后卵型鲳鲹冷藏期间的感官品质变化结果见图4。
由图4可知,接种3株优势腐败菌后的卵形鲳鲹感官分值总体呈下降趋势,而且变化规律基本一致,感官分值均低于对照组,新鲜的卵形鲳鲹切面色泽正常,富有光泽,肌肉组织纹理清晰,具有海水鱼固有的清香,富有弹性。但是随着贮藏时间的加长,冷藏卵形鲳鲹鱼体表面的色泽逐渐变暗淡,失去原来的光泽,变软且失去弹性,散发着腥臭味,肌肉纹理模糊,品质明显下降,逐渐趋向于腐败状态。到达第6 d时,对照组为5.6分<6分,超过可接受点,6 d时接种腐败希瓦氏菌的感官分值为3分,说明已经高度腐败。

图4 接种不同优势腐败菌的卵形鲳鲹冷藏期间感官品质的变化Fig.4 Sensory quality changes of Trachinotus ovatus inoculated with different dominant spoilage bacteria during chilled storage
2.5.2 菌落总数变化 菌落总数(TVC)的变化能够反映蛋白质和氨基酸分解代谢程度,Al-Daqal等[32]规定水产品可食用的菌落总数对数值上限是6.0,超出即为腐败。接种不同优势腐败菌后卵型鲳鲹冷藏期间的菌落总数变化结果见图5。
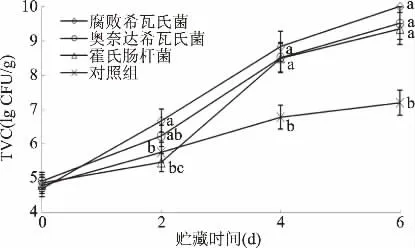
图5 接种不同优势腐败菌的卵形鲳鲹冷藏期间TVC值的变化Fig.5 TVC value changes of Trachinotus ovatus inoculated with different dominant spoilage bacteria during chilled storage
图5 显示,冷藏卵形鲳鲹接种优势菌后,随着贮藏时间的延长,菌落总数均呈增加趋势,对照组增加较为缓慢,贮藏第2 d后,除霍氏肠杆菌和对照组外,2株希瓦氏菌的菌落总数均已超过6.0,在4 d以后,3株菌株TVC的对数值均超过8.0,6 d时,腐败希瓦氏菌的TVC对数值达10,随着卵形鲳鲹鱼肉中的营养物质的消耗和有机酸、胺类物质代谢产物的积累,抑制了腐败菌生长[33],使得4 d后TVC的增加幅度逐渐减慢。
2.5.3 总挥发性盐基氮指标的测定 TVB-N值是评价水产品新鲜度的重要指标,样品鲜度分级标准参考GB/T 18108-2019[34],优级品TVB-N≤15 mg/100 g;合格品TVB-N≤30 mg/100 g,超过30 mg/100 g为腐败。接种不同优势腐败菌后卵型鲳鲹冷藏期间的总挥发性盐基氮值变化结果见图6。
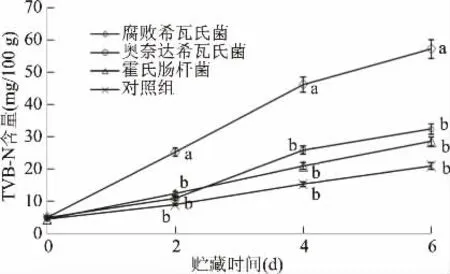
图6 接种不同优势腐败菌的卵形鲳鲹冷藏期间TVB-N值的变化Fig.6 TVB-N value changes of Trachinotus ovatus inoculated with different dominant spoilage bacteria during chilled storage
图6 可知,接种5、16号菌株和对照组的TVB-N值总体增加较为缓慢,接种2号腐败希瓦氏菌增加速度最快,第6 d时,其TVB-N值高达49.7 mg/100 g,远远高于5、16号菌株,说明此时卵形鲳鲹已经高度腐败,与感官评价结果比较一致。另外,不同腐败菌因营养要求不同,代谢也会存在一定差异,因此将菌落总数与挥发性盐基氮相结合,计算产量因子Y(TVB-N/CFU),可以定量表征各优势腐败菌的致腐能力。表4是腐败希瓦氏菌、奥奈达希瓦氏菌、霍氏肠杆菌接种无菌卵形鲳鲹鱼块后的产量因子测定结果。
由表4的TVB-N产量因子(YTVB-N)可以确定致腐能力大小排序为:腐败希瓦氏菌>奥奈达希瓦氏菌>霍氏肠杆菌;腐败希瓦氏菌的致腐因子为,值最大,表明其致腐能力最强;奥奈达希瓦氏菌和霍氏肠杆菌致腐因子接近,分别为3.92×10-8、3.11×10-8mg-1TVB-N/CFU,二者无显著性差异(P>0.05),因此将腐败希瓦氏菌确定为4℃冷藏卵形鲳鲹的优势腐败菌。腐败希瓦氏菌产量因子的致腐能力低于冷藏鲈鱼片[21]的腐败希瓦式菌属致腐能力;远高于罗非鱼[20]和冷藏鲢鱼[35]腐败希瓦氏菌的致腐能力;腐败希瓦氏菌致腐能力的不同,可能与水产品的所含营养成分的差异及代谢水平不同有关。
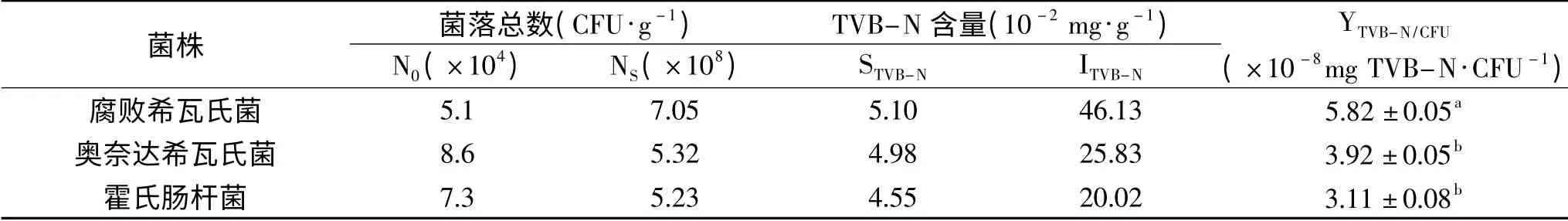
表4 3种优势腐败菌的TVB-N产量因子Table 4 TVB-N yield factors of 3 dominant spoilage bacteria
年益莹等[9]发现,酸化减菌处理金鲳鱼肉块微冻(-3℃)贮藏末期的优势菌为假单胞菌;张璟晶等[33]发现市售冰鲜银鲳的优势腐败菌是假单胞菌和希瓦氏菌;唐文静等[19]确定了冷藏海鲈鱼的优势腐败菌为假单胞菌、草莓假单胞菌和腐败希瓦氏菌;Gill等[36]研究认为假单胞菌属和希瓦氏菌属是冷链流通中高水分蛋白食品的特定腐败菌,Lone等[37]研究发现,在不同的贮藏和加工过程中水产品中的特定腐败菌是不同的,海水鱼冷藏过程中的SSO为希瓦氏菌属,淡水鱼冷藏过程中的SSO为假单胞菌属。如带鱼[38]、牙鲆[39]、鲈鱼[21]等水产品的特定腐败菌均为腐败希瓦氏菌。
本研究假单胞菌属在贮藏中期比例较高,后期基本消失,腐败希瓦氏菌后期占比最高达48%,且致腐能力又强,成为冷藏卵形鲳鲹的优势腐败菌。腐败希瓦氏菌为革兰氏阴性杆菌,其腐败活性强,能把氧化三甲胺还原成三甲胺,产生硫化氢等挥发性气体[40],导致水产品出现鱼腥恶臭味[5]。并能形成生物被膜使水产品表面发粘[41],表现出一系列腐败特性。腐败希瓦氏菌等为主的优势腐败菌数量及种类,与水产品货架期也存在显著关联[42]。因此腐败希瓦氏菌是导致低温贮藏水产品发生腐败的重要因素之一。
3 结论
本研究通过菌落形态、部分生理生化指标及16S rDNA分析,从4℃冷藏卵形鲳鲹鱼肉中获得16株腐败细菌,分别归属于6个属,假单胞菌属、葡萄球菌属、芽孢杆菌属、乳酸杆菌属等大多出现在初期和中期;希瓦氏菌属种类最多,其中腐败希瓦氏菌和奥奈达希瓦氏菌、肠杆菌属的霍氏肠杆菌在贮藏后期占比较高;对贮藏期后期的3株数量优势菌进行致腐能力定量测定发现,腐败希瓦氏菌的致腐能力远高于奥奈达希瓦氏菌和霍氏肠杆菌,最终确定腐败希瓦氏菌(Shewanella putrefaciens)是4℃冷藏卵形鲳鲹的优势腐败菌;该研究可为后续靶向抑菌延长冷藏卵形鲳鲹货架期提供理论参考。
