植物中天然橡胶合成及研究进展
2021-06-22李海英马春泉
杨 超,李海英,马春泉,*
(黑龙江大学 a.农业微生物技术教育部工程研究中心;b.生命科学学院,哈尔滨 150080)
0 引 言
天然橡胶是基础工业原料之一,广泛应用于航空航天、医疗器械、交通运输等军用和民用工业,在诸多高精度领域中均无法被合成橡胶所取代。天然橡胶是由异戊二烯聚合生成的一种生物高分子材料(C5H8)n,包括顺式异戊二烯(cis-polyisoprene,CPI)和反式异戊二烯(trans-polyisoprene,TPI)两种同分异构体,简称为顺式胶和反式胶[1]。顺式胶具有高弹性、耐冲击、应诱变结构特征、耐穿刺、耐撕裂等诸多优良的理化性能,在低温下也能保持优秀的弹性和延展性[2-3],顺式胶的这些优异特性与其自身的网状结构、高分子量以及胶乳(橡胶颗粒的胶状悬浮物)中的一些次要成分(如蛋白质、脂类、碳水化合物和矿物质)息息相关。与顺式胶相比,反式胶的综合性能稍差,但其优势在于硬度高、热膨胀/收缩系数较低、绝缘性好以及耐酸碱腐蚀[4]。以化石资源为原料生产的合成橡胶虽为反式胶,但目前技术手段生产出的合成橡胶中含有的其他分子结构杂质,破坏了合成橡胶中的大分子排列规律,影响了其理化性能,致使工业未能生产出性能上与天然橡胶相匹配的合成橡胶材料。此外,合成橡胶来源的石油供应和价格的不确定性以及快速增长的经济体对橡胶的高需求均导致了天然橡胶和合成橡胶价格的迅速上涨,提高了开发新型天然橡胶替代资源的必要性与紧迫性。因此,寻找和开发与天然橡胶相近的产胶替代植物是当前的研究热点之一。
高分子量是决定天然橡胶质量的重要因素之一。目前在自然界中已经鉴定出不少于8科、300属、2 500 多种可以产生顺式胶的植物中[5],除了巴西橡胶树(Heveabrasiliensis)外,仅有橡胶草(Taraxacumkok-saghyz)和银胶菊(Partheniumargentatum)两个物种可以合成产量可观且分子量高的顺式胶;自然界中可生产反式胶的植物数量也较少,目前仅有杜仲(Eucommiaulmoides)报道较为广泛。无论是生产顺式胶还是反式胶,理想的产胶植物需要具备生长快、产胶量高、适应范围广等优良特性,使其推广到农作系统中,并且根据市场需求考虑与其他作物进行轮作,以更好地满足人类生活的需要。
1 天然产胶植物
1.1 巴西橡胶树
巴西橡胶树(Heveabrasiliensis)是目前天然橡胶唯一的商业来源。巴西橡胶树的生长周期长,6~7 a才能进入出产期,给巴西橡胶树遗传育种带来了困难[6]。同时,巴西橡胶树对生长环境要求十分严格,种植条件限制了其只能在热带环境下种植,也成为了巴西橡胶树的种植无法扩大到非热带国家的主要障碍。此外,现如今对巴西橡胶树威胁最大的疾病是南美叶疫病(SALB),是由南美叶疫病菌(Microcyclusulei)引起的真菌性疾病[7]。由于东南亚地区主要引进并推广种植的均为Henry Wickham系统22株橡胶树品系,品种单一化导致该地区橡胶树遗传基因狭窄,生产系统的局限性和脆弱性使其更容易受南美叶疫病(SALB)的威胁,给通过基因工程手段改造培育高产抗病橡胶树品种带来了困难[8]。开发天然橡胶替代植物来源的另一原因是橡胶树胶乳中的蛋白质可引起人体的过敏反应,而且对于乳胶过敏反应的人群数量约占普通人群的1%~6%,研究表明,医护人员中存在过敏反应风险高达17%[5]。
1.2 橡胶草
橡胶草(Taraxacumkok-saghyz)是一种草本植物,是前苏联科学家Rodin于1931—1932年在哈萨克斯坦境内天山山谷中发现并引种培育,根是主要产胶部位,橡胶草橡胶的分子量非常高,在质量方面与巴西橡胶树橡胶相似,在二战期间橡胶草橡胶被广泛应用[9]。橡胶草作为巴西橡胶树最具竞争力的产胶替代植物,具有众多优点:①橡胶草适应能力强,可在温带地区广泛种植,且具有良好的耐盐碱能力;②橡胶草产胶周期短,仅一年就可收获,在选育高产胶量品种方面具有优势;③橡胶草目前已建立较为成熟的再生及遗传转化体系,可利用分子育种的手段培育橡胶草新品种;④橡胶草在田间可与其它作物轮流种植或者间隔种植,充分利用土地面积,增加经济效益;⑤橡胶草具有较强的经济竞争力,加工的副产品菊糖可应用于食品和非食品领域,剩下的植物残渣可以发酵成生物乙醇和沼气[10]。
尽管橡胶草拥有和橡胶树一样的乳管,但却无法通过割胶来收获胶乳,只能通过破碎橡胶草的根后才能提取;而且橡胶草橡胶颗粒相比巴西橡胶树含有更多的蛋白质,致敏可能性更高,这也增加了人体对橡胶草橡胶过敏的风险,因此橡胶草需要考虑应用于传统、非医疗用途,如轮胎制造等[11]。另外,橡胶草基本上未经驯化,存在一些固有问题,如发芽期间需要稳定的含水量、生长速度慢、与杂草的竞争力差、橡胶产量只有在成熟时才能测量、高度杂合度以及自交不亲和性等特性,这些问题严重限制了橡胶草的替代和应用速度,也成为了正在重点攻克的研究领域[10]。因此,在未来橡胶草成为商业上可行且具有竞争力的天然橡胶植物之前,通过分子育种和常规育种相结合,提高橡胶草橡胶产量和改善农艺性状至关重要,最终目标是培育出地下生物量高、橡胶积累量高的新品种。
1.3 银胶菊
银胶菊(Partheniumargentatum),生长于墨西哥和美国南部半干旱地区的一种多年生灌木。银胶菊需要1 000~2 200 mm的总用水量(灌溉加降雨)才能保证橡胶的产量。银胶菊橡胶分子量与巴西橡胶树橡胶基本相同,因而也是主要的产胶替代植物之一[12]。
银胶菊从种植到收获和胶乳生产可以完全机械化,但银胶菊胶乳在树皮薄壁细胞中形成毫米大小的颗粒,必须通过机械手段彻底破坏植物材料,从单个细胞中释放出胶乳,再通过离心和乳化等步骤来纯化胶乳,或者通过溶剂萃取来分离橡胶,因而银胶菊橡胶加工技术十分复杂,增加了生产成本[13]。银胶菊橡胶的优势在于它所产橡胶的蛋白质含量比巴西橡胶树橡胶低得多,相关蛋白数量不超过10种,因此银胶菊橡胶满足了市场上对低过敏性橡胶的需求,如医用手套、导管等高附加值的商业产品。但银胶菊的缺点也给其商业化进程带来了障碍。在欧洲和美国大部分地区,银胶菊只有部分品种驯化成两年或多年生植物,而在中国,银胶菊被归为外来入侵物种,存在着破坏生态环境的风险。此外,银胶菊不耐冷,只有在特定的冬季温度下才能生产高质量的橡胶,这也限制了银胶菊橡胶的产量。
1.4 杜仲
杜仲(Eucommiaulmoides),分布于我国温带和亚热带地区,引种栽培面积达35×104hm2。杜仲作为我国本土生长的特有树种,是珍贵的中药材,其组织器官包括树皮、叶片和种子均可入药,杜仲的这些部位均分布有高分子量的反式胶,是具有开发潜力的橡胶资源[14]。
杜仲的生态环境适应能力极强,能够在酸性土壤和盐碱土壤中生长,可在我国大部分地区种植,开发前景巨大。另外,杜仲胶具有橡塑二重性,即在常温下呈现类似塑料的状态,而在50 ℃以上表现为橡胶的性质,是具备热塑性及热弹性的优良功能材料[15]。这一性质使得杜仲胶在航天、船舶等领域得到应用,但也极大地限制了其在常温下的应用范围。目前,对于杜仲的研究主要局限于药理研究,对杜仲胶合成及提取的研究相对较少,制约了杜仲胶在国内的发展。生物量多的叶片是提取杜仲橡胶的主要原料,但含胶量仅2%~6%,提取过程中需要处理大量废渣,提高了杜仲胶提取的加工生产成本[16]。
2 天然橡胶的生物合成机制
天然橡胶的生物合成途径(图1)包括3个阶段:①异戊烯基焦磷酸(IPP)的形成;②聚异戊二烯基焦磷酸链的延伸;③橡胶分子的合成[5]。
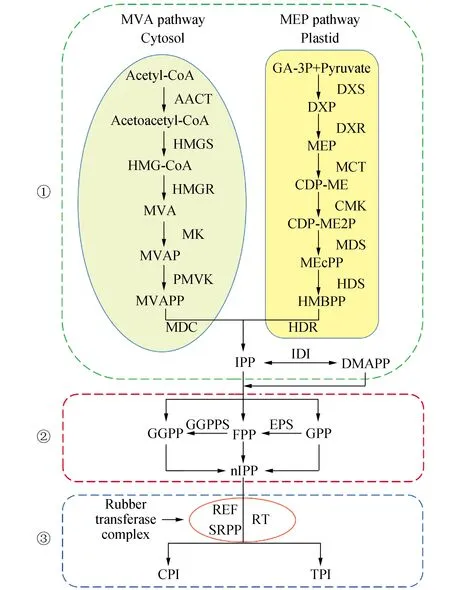
图1 天然橡胶的生物合成途径Fig.1 Biosynthesis pathway of natural rubber
2.1 异戊烯基焦磷酸(IPP)的合成
异戊烯基焦磷酸(IPP)是橡胶生物合成的单体亚单位,由乙酰辅酶A或3-磷酸甘油醛和丙酮酸通过两种途径合成,即甲羟戊酸途径(MVA)和甲基赤藓糖醇磷酸盐途径(MEP),这两个途径分别在细胞质和质体中进行[1,8]。MVA途径也称作细胞质途径。天然产胶植物体内的蔗糖由转化酶催化下分解为葡萄糖和果糖,然后葡萄糖通过糖酵解途径转化为丙酮酸,再由丙酮酸脱氢酶复合体生成乙酰辅酶A,之后乙酰辅酶A作为起始底物进入MVA途径,经过6步的缩合反应生成IPP[5]。MEP途径也称作质体途径。MEP途径是以糖酵解途径产生的丙酮酸和甘油醛-3-磷酸为起始底物,经过7步反应合成IPP和二甲基烯丙基焦磷酸(DMAPP)[8]。
2.2 聚异戊烯基焦磷酸链的延伸
DMAPP是IPP的双键异构体,两者可通过异戊烯基焦磷酸异构酶(IDI)相互转化,DMAPP通过反式异戊烯基转移酶(TPT)与IPP进行连续顺式-1,4-缩合反应,形成牻牛儿基焦磷酸(GPP)、法尼基焦磷酸(FPP)和牻牛儿基牻牛儿基焦磷酸(GGPP)。上述这些反式短链异戊二烯基焦磷酸已被证明可以作为橡胶合成的引发剂,引发剂的分子量与天然橡胶的合成速度呈正相关[6]。异戊二烯基焦磷酸与IPP结合,引发聚异戊烯基焦磷酸链的结合,这一过程需要二价金属阳离子(Mg2+或Mn2+)的参与,之后的延伸过程就是异戊烯基焦磷酸在异戊烯基转移酶(PT)的催化下不断地连接到聚异戊烯基焦磷酸焦磷酸长链上[7]。
2.3 橡胶分子的合成
橡胶分子的合成是聚异戊二烯链从橡胶转移酶复合体上解离下来的过程,之后橡胶分子就会贮存于橡胶粒子内部。橡胶分子终止合成的信号仍需要进一步探讨,有观点认为聚异戊二烯链的终止可能涉及到由焦磷酸酶催化焦磷酸基团解离而形成羟基这一过程,也有观点认为由于橡胶转移酶(RT)附着在橡胶粒子的膜上,因此在合成橡胶分子时,会产生一定程度的几何空间阻碍而终止合成橡胶[8]。
3 天然橡胶生物合成相关基因和关键酶的研究
3.1 羟甲基戊二酰辅酶A还原酶(HMGR)
3-羟基-3-甲基戊二酰CoA(HMG-CoA)在HMGR的催化下生成甲羟戊酸(MVA),此过程不可逆。像脱落酸、赤霉素、类胡萝卜素以及辅酶Q等初生代谢物的前体就是MVA[14],HMGR是MVA途径的一个限速酶。在橡胶树中发现了3个HMGR基因(HMGR1-3),其中HMGR1在乳管中表达量最高,可以被乙烯诱导,与天然橡胶合成最为相关,烟草中异源表达HbHMGR1后发现处理组HMGR活性与野生型相比增加了4~8倍,甾醇含量也增加到6倍[17]。从构巢曲霉(Aspergillusnidulans)克隆得到的AnHMGR基因通过农杆菌介导转化银胶菊,转基因银胶菊与野生型银胶菊相比,胶乳产量显著增加,天然橡胶含量增加了65%[12]。
3.2 1-脱氧木酮糖-5-磷酸合成酶(DXS)
DXS是MEP途径中的首个关键酶,丙酮酸和3-磷酸甘油醛在其催化下生成脱氧木酮糖磷酸(DXP),这一过程需要焦磷酸硫胺素(TPP)作为辅酶参与。DXS基因最早在大肠杆菌(Escherichiacoli)中被分离,同时基因敲除结果发现表达DXS基因菌株的萜类化合物产量显著提高[18]。拟南芥(Arabidopsisthaliana)是最早分离出DXS基因的植物之一,将该基因转入到薰衣草中异源表达,显著提高了转基因薰衣草中主要成分单萜烯精油的含量[19]。
3.3 1-脱氧-D-木酮糖-5-磷酸还原异构酶(DXR)
DXR是MEP途径中的第二个关键酶,参与催化DXP生成MEP,该过程需要二价金属离子(Mg2+、Mn2+或Co2+)和NADPH作为辅酶参与。药用植物雷公藤(Tripterygiumwilfordii)过表达DXR基因植株的雷公藤内酯醇的产量提高,药用价值增加[20]。DXR过表达拟南芥植株中类胡萝卜素的含量也有明显提高[21]。
3.4 法尼基焦磷酸合酶(FPS)
法尼基焦磷酸合酶(FPS)是天然橡胶合成途径中限速酶之一,催化两分子的IPP和一分子的DMAPP以1~4头尾结合的方式连续缩合形成法尼基焦磷酸(FPP)。法尼基焦磷酸是植物体内多种萜类衍生物的前体,参与合成的物质有类胡萝卜素、甾醇、质体醌和倍半萜衍生物(如植保素)等对植物生长发育及抗病过程具有重要作用。Cornish K[22]研究发现,天然橡胶的生物合成速率和FPP浓度呈正相关,即细胞内FPP浓度越高,天然橡胶的生物合成速率就越快。因此,可以通过分子生物学技术过表达FPS,增加植物细胞内FPP的浓度,从而实现提高天然橡胶产量的目标。
3.5 牻牛儿基牻牛儿基焦磷酸合酶(GGPPS)
GGPPS在植物体内催化合成萜类物质合成前体之一的牻牛儿基牻牛儿基焦磷酸(GGPP)。最早在辣椒(Capsicumannuum)中克隆出GGPPS基因,氨基酸序列比对表明GGPPS包括7个保守的氨基酸残基结构域。GGPPS过表达橡胶草株系的根部胶乳含量明显提高,同时外源施加茉莉酸甲酯(MeJA)使GGPPS的表达量极显著提高[10]。
3.6 橡胶延长因子(REF)
橡胶延长因子(REF)是一种橡胶粒子膜蛋白,分子量为14.6 kDa。研究分析表明REF和SRPP(小橡胶粒子蛋白)是来自同一原始基因的同源蛋白,它们属于一个与植物胁迫相关的蛋白家族,即REF/SRPP超家族,包含REF和SRPP两个亚家族。REF在对杜仲叶片、茎皮研究发现,EuREF1的表达量与含胶量呈正相关[16]。
3.7 小橡胶粒子蛋白(SRPP)
橡胶生物合成在乳管细胞内的橡胶粒子中进行,小橡胶粒子蛋白(SRPP)在橡胶生物合成中发挥重要的作用,是决定橡胶生物合成速率和胶乳产量与橡胶质量的关键橡胶粒子蛋白。SRPP定位在小橡胶粒子膜上,分子量约为24 kDa,ORF全长612 bp,SRPP与REF具有较高同源性,而REF则已被证实参与天然橡胶的生物合成。Xie Q等[11]在橡胶草成熟根中橡胶合成相关基因表达的研究结果显示,SRPP极可能通过构建一个最有利且稳定的橡胶粒子体系而影响橡胶的生物合成和最终存储,进而在橡胶合成过程中发挥着至关重要作用。银胶菊中也发现SRPP能促使异戊二烯添加到橡胶分子中[13]。
3.8 异戊烯基转移酶(PT)
IPP与DMAPP通过缩合反应生成GPP,以此为基础连续加入IPP的链伸长反应由PT催化合成。PT又称为异戊烯基二磷酸合酶,根据产物双键构型可分为催化反式双键的E-PT和催化合成顺式双键的Z-PT。杜仲中FPS作为E-PT参与合成天然反式橡胶[23]。橡胶转移酶(RT)是PT中的一员,其酶活性影响着天然顺式橡胶的质量和产量,是产胶能力评价指标之一。目前在巴西橡胶树中发现2个高度同源的橡胶转移酶RT1和RT2,这两个酶在胶乳中特异性表达[24],同时Schmidt T等[9]研究发现橡胶草中有RT参与橡胶合成,其活性与橡胶产量呈正相关。
4 结论与展望
天然橡胶的生物合成过程复杂,至今仍不完全清楚橡胶在橡胶树中确切的生物学功能。虽然胶乳可起防卫作用,但橡胶分子本身并无这种功能,并且橡胶分子在植物中似乎属于代谢终产物,迄今还未发现可降解此类分子的具体酶类。尽管已知天然产胶植物中大多数植物的天然橡胶生产量少、质量差、分子量低,但随着对天然橡胶生物合成途径研究的不断深入以及基因工程技术的不断进步,天然产胶植物的筛选及改良必将集中开发高质量产胶植物的培育上,可缩短天然橡胶的产胶周期。此外,充分利用当下成熟的基因编辑技术在传统植物体内引入编码天然橡胶生物合成关键酶基因,或者进行天然橡胶合成途径整体转移的合成生物学技术的开发与利用,将成为未来天然产胶植物研究的热点,这些策略需要建立在植物中天然橡胶产量和质量的生物化学调控具有一个全面深入的了解基础之上。
