坡度对亚高寒草甸狼毒群落植物多样性的影响
2021-06-18崔博亮安彦明张起鹏陈克龙
崔博亮,安彦明,张起鹏,蒲 萌,陈克龙
(1.青海师范大学地理科学学院,青海西宁810008;2.十堰市车城高级中学,湖北 十堰442000;3.聊城大学环境与规划学院,山东聊城 252000;4.甘肃民族师范学院,甘肃 合作747000;5.青藏高原地表过程与生态保育教育部重点实验室 /青海省自然地理与环境过程重点实验室,青海 西宁810008)
青藏高原被称作“世界第三极”,是全球海拔最高,面积最大的陆地生态系统[1]。亚高寒草甸是青藏高原分布较为广泛的陆地生态系统,是青藏高原特色牧业生产基地和发挥生态屏障的重要载体[2-3],在高原草业、畜牧业中占有十分重要的地位,对亚高寒草甸可持续利用的研究直接关系到当地牧民的生存与发展[4]。然而,在自然因素和人为因素的作用下,亚高寒草甸天然草地生产能力下降,狼毒等毒杂草群落面积不断增多,占据了优良牧草生存所必需的光、热和水等资源,导致天然草原质量下降,严重制约和影响了天然草原畜牧业的发展和高寒草地的生态功能[5]。
瑞香狼毒(Stellera chamaejasme)为瑞香科狼毒属多年生草本植物[6],其地理分布主要在我国东北、华北、西北、西南、内蒙古及青藏高原等海拔较高、气候恶劣的干燥向阳山坡及高山草地地带[7],是我国退化草地上广泛分布且最为常见的有毒植物之一[8]。狼毒对环境具有极强的适应和改造能力[9],它的侵蚀和蔓延可降低天然草原草地质量和草原生产力,导致草地严重退化,是我国天然草原生物多样性维持和畜牧业发展的安全隐患[10]。一般情况下,在适度放牧草地上,瑞香狼毒作为常见的伴生种存在,而在过度放牧出现退化的草地常以优势种存在[11],作为天然草原退化的一种标志性植物,是草地群落逆向演替的结果,能形成以狼毒为优势种的毒草退化型草地景观[12]。因此,开展退化草地狼毒等毒杂草群落的研究具有重要的经济社会和生态意义。目前,国内外学者多采用传统地面及地下取样调查的方法来研究退化草地中的狼毒,一定程度上丰富了狼毒种群的相关信息[13]。主要研究有不同草地管理措施对狼毒型退化草地群落结构及牧草品质的影响[14];狼毒种群数量对青海湖环湖草地土壤肥力的影响[15],毒杂草退化草地植被群落特征的研究[16],退化草地狼毒群落土壤种子库的空间格局分析[17],以及退化草原狼毒个体年龄判定方法及其种群年龄结构的研究[18]等。但是在高寒地区考虑到坡度因子并结合物种功能群划分和物种多样性来分析退化草地狼毒群落在亚高寒草甸的相关研究较少。鉴于此,本研究以青藏高原东北部边缘甘南亚高寒草甸草地群落为研究对象,采用样方调查方法,通过对不同坡度狼毒群落的分布现状、特征、多样性和环境条件等方面的研究,来探讨狼毒群落对该区域草原退化程度的指示作用[19],揭示不同坡度上狼毒群落植被的分布格局以及其对生境和微气候的适应策略[20],以期为该地区生态环境评估及政府管理部门制定狼毒防除措施及生态保护和管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于甘南藏族自治州合作市当周草原和 扎 油 沟 地 带(102°53′10″–102°53′26″E,34°57′00″–34°57′59″N;102°54′00″–102°54′11″E,32°00′11″–35°00′07″N,海拔2976~3062 m),高原大陆性季风气候特征明显,冷季漫长,平均气温2℃,最冷月(12月−2月)平均气温−8.9℃,极端最低气温− 24℃。最热月(6月−8月)平均气温11.5℃,极端最高气温28℃,0℃以上年积温1730℃·d,年均降水量560 mm,50%集中在6月− 8月,雨热同期。年蒸发量1222 mm,平均无霜期48 d,主要自然灾害为霜冻、冰雹和阴雨[21]。土壤类型为高山草甸土,植被条件良好,阴坡主要植物为嵩草(Kobresia myosuroides)、矮针蔺(Trichophorum pumilum)等莎草科植物以及异针茅(Stipa aliena)、喜马拉雅早熟禾(Poa himalayana)、垂穗披碱草(Elymus nutans)等禾本科植物;而阳坡主要植物为金露梅(Potentilla fruticosa)、嵩草、垂穗披碱草及多种杂类草。研究区域为冬季牧场,中等放牧强度,试验期间不受放牧活动干扰。
1.2 试验设计与样地调查
调查于2018年6月− 7月进行,该时段是研究区一年当中植物生长最为旺盛、草地生产力最高的月份。在研究区年际放牧强度一致、所处环境条件相近的草地阳坡选取狼毒群落类型分布较为典型的地段,采用样方法进行调查。为更好地对样地的总盖度和所有物种的分盖度、高度和密度进行测定记录,本研究根据国际地理学联合会地貌调查与地貌制图委员会关于地貌详图应用的坡地分类方法并结合调查样地实际情况,将调查样地坡度分为Ⅰ(0°~5°]、Ⅱ(5°~15°]、Ⅲ(15~25°]、Ⅳ(25°~35°]及Ⅴ(> 35°)5个坡度等级[22],在每个坡度的同一海拔高度上选取4~8个样方(草本样方面积为0.5 m ×0.5 m,灌丛样方面积为2 m×2 m)进行草地群落调查,对每个样方的植物种类、个数、高度、盖度、多度及频度等指标进行调查记录,除以上指标外,还记录每一个样方的海拔、经纬度、土壤、植被类型及干扰状况等环境指标,共调查记录了27个样方群落。调查过程中参考张起鹏[23]的调查方法,不同类型植物采取不同的调查计算方法,对于不可再分的密丛型物种以有主茎的植物为一株,匍匐茎型植物按照其主根计算,调查过程中尚不能识别的植物将其采集标本并编号后带回实验室鉴定种属,无法鉴定到种的植物鉴定到属。结合现场调查和数据统计分析发现,研究区狼毒主要分布在当周草原和扎油沟等地区(图1)。
1.3 重要值与功能群划分
重要值是衡量某个物种在所在群落中重要性的综合数量指标[24-25],本研究利用重要值大小来确定样地内物种的优势度,采用相对高度、相对盖度和相对多度的平均值计算各物种的重要值(IV),其计算公式[26]为:

式中:IV为物种的重要值;RH为物种相对高度;RC为物种相对盖度;RD为物种相对多度。
相对高度=(样方内某物种的平均高度/样方内所有物种的平均高度之和)×100%;
相对盖度=(样方内某物种的盖度/样方内所有物种盖度之和)×100%;
相对多度=(样方内某物种的多度/样方内所有物种的多度之和)×100%;
相对频度=(样方内某个物种的频度/样方内所有物种的频度之和)×100%[27-28]。
本研究结合研究区高寒植物群落物种组成及其特点,参照文献[29-30]中对植物功能群的划分方法,将研究区功能群划分为莎草科植物、多年生根茎禾草、多年生丛生禾草、灌木与半灌木、多年生杂类草以及一、二年生草本植物6类。
1.4 多样性测算
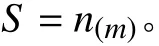
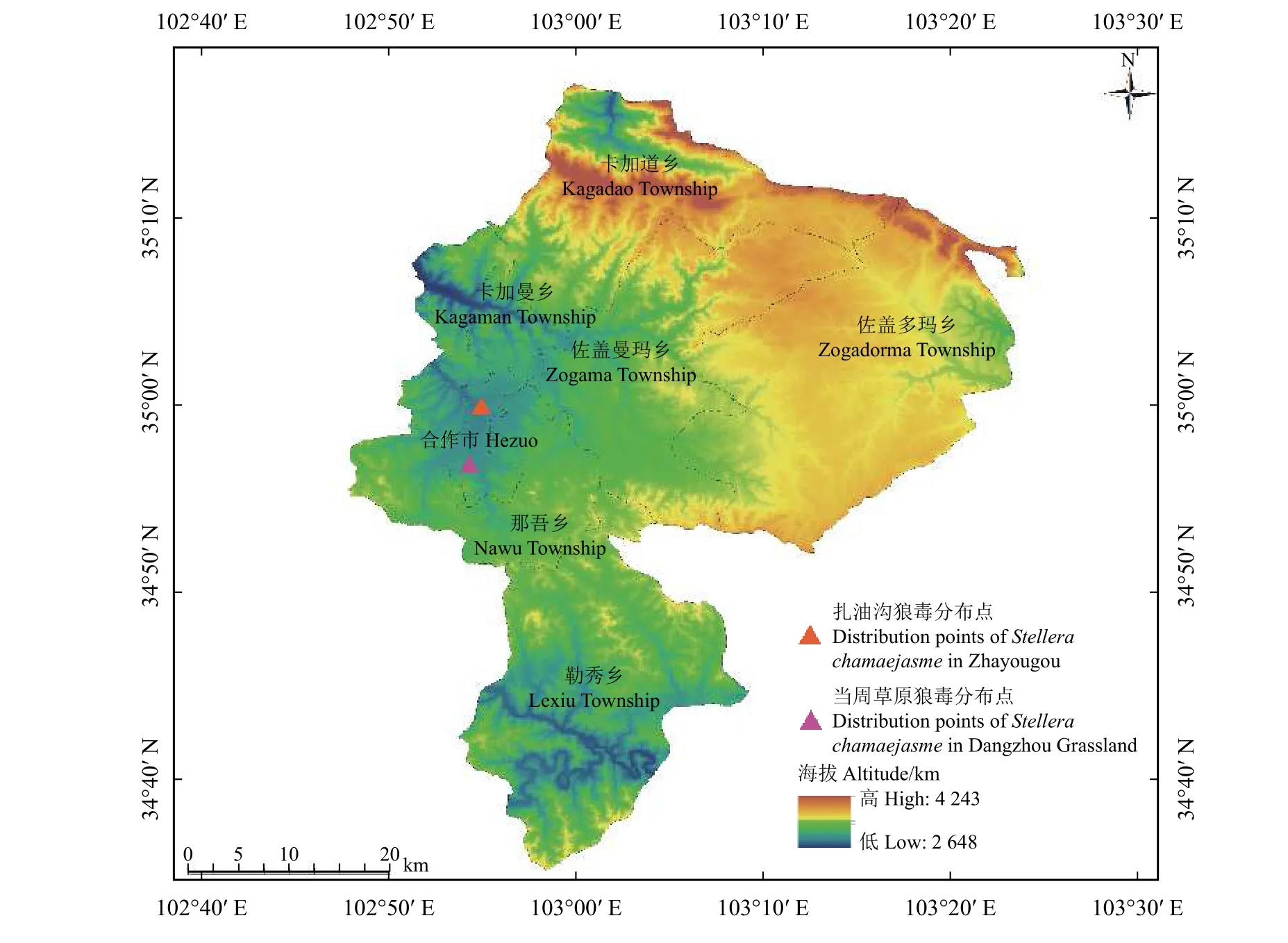
图1 调查样地狼毒分布位置图Figure 1 Distribution map of Stellera chamaejasme in the survey area
式中:S表示物种丰富度指数;n表示调查样方内的物种数目;m表示调查样方的面积值[31]。

式中:H′表示Shannon-Wiener多样性指数;Pi表示物种i的重要值占所有个体重要值的比重;IVi表示第i个物种的重要值。

式中:J表示Pielou均匀度指数;H′max表示在最大均匀条件下的群落多样性指数[31]。
1.5 数据分析
所有原始数据均采用SPSS 22.0进行数据统计分析,采用Excel 2010制图。
2 结果分析
2.1 不同坡度亚高寒草甸狼毒植物群落组成
在27个狼毒群落调查样地中,共记录到植物约50种,主要有嵩草、牡蒿(Artemisia japonica)、蒲公英(Taraxacum mongolicum)、冷蒿(A. frigida)、猪毛蒿(A. scoparia)、麻花头(Serratula chinensis)、薄雪火绒草(Leontopodium japonicum)、蕨麻(Potentilla anserina)、乳白香青(Anaphalis lactea)、垂穗鹅观草(Roegneria nutans)、扁蕾(Gentianopsis barbata)、黄花棘豆(Oxytropis ochrocephala)、匍匐栒子(Cotoneaster adpressus)、垂穗披碱草、狼毒、异针茅、矮针蔺、金露梅等物种。根据样地中植物出现的频率,调查区排名前十的植物物种分别为嵩草、黄花棘豆、狼毒、冰草、喜马拉雅早熟禾、蕨麻、牡蒿、蒿子、薄雪火绒草、乳白香青;其频度分别为23.6%、23%、17.7%、13.7%、10.9%、10.5%、7%、6.7%、5.82%、4.7%。在坡度Ⅳ人畜活动较为频繁地带,植物群落较为丰富;坡度Ⅴ地带由于坡度升高,地势较为陡峭,土壤质地疏松,调查样方中物种数有开始减少的趋势,且出现部分裸露地。不同坡度上的主要物种组成见表1。

表1 不同坡度上主要物种组成Table 1 Main species composition on different slopes
2.2 不同坡度亚高寒草甸植物群落功能群分析
莎草科植物主要由嵩草构成,占植物总数的2.0%,优势度为23.6%,在不同坡度上,莎草科植物重要值的分布为Ⅱ>Ⅳ> Ⅰ>Ⅴ> Ⅲ,坡度Ⅱ重要值最高,为25.1%,坡度Ⅲ最低,为14.6%,变化幅度为10.5%,其中坡度Ⅲ和坡度Ⅴ重要值变化幅度不是很明显,仅有1.0%(图2);多年生根茎禾草由冰草、喜马拉雅早熟禾等植物组成,占总物种数的4.1%,优势度为16.3%,在不同坡度上其重要值的分布为Ⅴ> Ⅳ> Ⅲ>Ⅰ> Ⅱ,除坡度Ⅱ重要值略低于坡度Ⅰ外,多年生根茎禾草重要值在其他坡度都呈现出随着坡度的升高而逐渐递增的趋势,其中,坡度Ⅱ重要值最低,为11.6%,坡度Ⅲ重要值最高,为28.4%,最高值和最低值的变化幅度为16.8%;多年生丛生禾草由异针茅构成,占总物种数的6.1%,优势度为25.0%,在不同坡度上重要值分布为Ⅴ> Ⅳ>Ⅰ> Ⅲ> Ⅱ,其重要值大致呈现出随着坡度升高而增大的变化趋势,其中坡度Ⅱ重要值最低,为7.2%,坡度Ⅴ重要值最高,为24.1%,最高值和最低值的变化幅度为16.9%;灌木与半灌木植物主要由金露梅构成,占总物种数的2.0%,优势度为5.7%,在不同坡度上其重要值分布为Ⅳ> Ⅴ>Ⅲ> Ⅱ>Ⅰ,灌木与半灌木在坡度Ⅰ上未发现相关物种,而在Ⅱ~Ⅴ坡度均有存在,呈现出先升高再降低的变化趋势,最高值出现在坡度Ⅳ,为14.9%,最低值在坡度Ⅱ,为4.8%,最高值和最低值的变化幅度为10.1%;多年生杂类草由狼毒、薄雪火绒草、乳白香青和珠芽蓼(Polygonum viviparum)等植物组成,占总物种数的73.6%,优势度为6.3%,在不同坡度上重要值分布为Ⅲ> Ⅱ> Ⅳ> Ⅴ> Ⅰ,整体呈现出先升高再降低的单峰变化趋势,在坡度Ⅲ达到峰值后降低,其中坡度Ⅰ重要值最低,为3.4%,坡度Ⅲ重要值最高,为14.8%,最高值和最低值的变化幅度为11.4%;一、二年生植物常见的植物有苦苣菜(Sonchus oleraceus)、葛缕子(Carum carvi)和泽漆(Euphorbia helioscopia)等,占总物种数的12.2%,优势度为4.4%,在不同坡度上的重要值分布为Ⅴ> Ⅳ>Ⅲ > Ⅱ> Ⅰ,重要值随着坡度的升高而逐渐增大,其中坡度Ⅰ重要值最低,为2.6%,坡度Ⅴ重要值最高,为25.8%,变化幅度为23.2%。
狼毒群落各类植物功能群重要值的最高值和最低值在不同坡度表现出不同的分布特征。各功能群最高值坡度分布为:莎草科植物在坡度Ⅱ,灌木与半灌木在坡度Ⅳ,多年生杂类草在坡度Ⅲ,多年生根茎禾草、多年生丛生禾草和一、二年生植物均在坡度Ⅴ。各功能群最低值坡度分布为:莎草科植物在坡度Ⅲ,多年生杂类草和一、二年生植物在坡度Ⅰ,多年生根茎禾草、多年生丛生禾草、灌木与半灌木在坡度Ⅱ。不同植物功能群对坡度等地形因子有着不同的选择差异[32],总体来看,本研究区大多数植物功能群在坡度Ⅳ和坡度Ⅴ上重要值最高,在坡度Ⅰ和坡度Ⅱ上重要值最低。
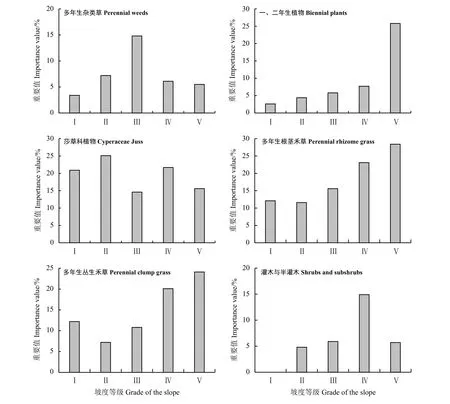
图2 不同坡度上狼毒群落植物功能群组成及其重要值Figure 2 Composition of plant functional groups and their importance value in Stellera chamaejasme community on different slopes
2.3 不同坡度亚高寒草甸狼毒群落多样性对比分析
在不同坡度上物种丰富度大小依次为Ⅳ> Ⅰ>Ⅱ> Ⅲ> Ⅴ(图3),在5个坡度中物种最丰富的是坡度Ⅳ,该坡度水热条件相对适宜,植被覆盖度较高,物种种类最为丰富,优势种为狼毒且分布较广,杂类草优势度较高;物种最少的是坡度Ⅴ,地势高且多不平坦,地表裸露度大,植被多样性逐渐减少,多杂草,优势种为禾本科植物和灌丛类,兼以根系发达的杂类草;狼毒分布最广的是坡度Ⅲ,其次是坡度Ⅱ。不同坡度上Shannon-Wiener多样性指数分布大小为Ⅳ>Ⅴ> Ⅰ>Ⅱ> Ⅲ,多样性分布最优的是坡度Ⅳ,该调查样地狼毒群落结构复杂,植物种类较为丰富,莎草科植物、禾本科植物、灌丛类及狼毒等多年生杂类草均有分布,生物多样性相对于其他生境群落要高;植物多样性最少的是坡度Ⅲ,物种以狼毒等杂类草为主,究其原因是该坡度人畜活动较为频繁,草地出现退化现象。在不同坡度上Pielou均匀度指数依次为Ⅳ>Ⅴ> Ⅰ > Ⅱ> Ⅲ,除Ⅳ、Ⅴ坡度物种均匀度指数较高外,Ⅰ、Ⅱ、Ⅲ坡度物种均匀度指数都较为均匀。
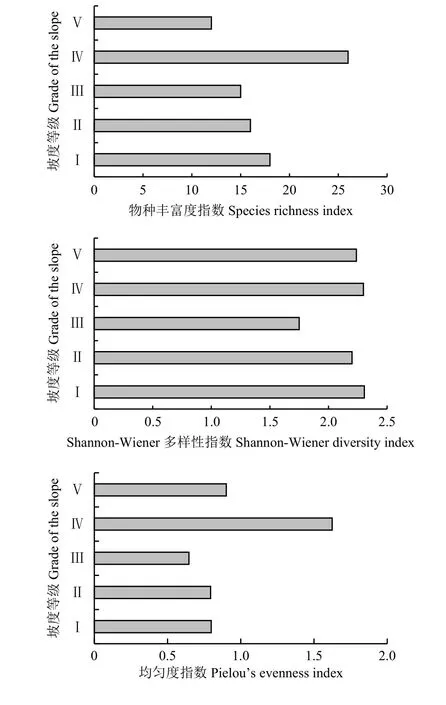
图3 不同坡度狼毒群落物种多样性指数Figure3 Speciesdiversity index of Stellera chamaejasme community on different slopes
总体来看,在Ⅰ坡度中,地势较低,利于积水,地形平坦且土层较厚,土壤湿润,植被繁茂,无狼毒分布,最高处群落总盖度可达100%,物种多样性和丰富度可观,优势种多为莎草科、禾本科植物等,植株高度相对较低、适口性好的优质牧草生长较好;在Ⅱ坡度中,海拔相对升高,坡度增大,但地势较低,水分条件好,植被覆盖度较高,物种种类较为丰富,优势种主要有莎草科、禾本科植物等物种,杂类草优势度有所上涨并出现狼毒分布;在Ⅲ坡度中,土壤养分及水分状况较好,狼毒分布较广且以建群种存在,多样性相比Ⅱ坡度减少,人畜活动较频繁,一些地方出现明显草地退化现象;在Ⅳ坡度中,坡度逐渐增大,海拔也进一步升高,该坡度水热条件适宜,植被覆盖度较好,物种种类最为丰富,优势种为狼毒且分布较广,杂类草优势度较高;Ⅴ坡度地势高,多不平坦,地表裸露度大,植被多样性逐渐减少,多杂类草,优势种为禾本科植物及灌丛,兼以根系发达的杂类草。
3 讨论
瑞香狼毒是我国天然草地上较为常见的有毒物种之一,在我国东北、华北、西北、西南及青藏地区的草地上都有不同程度的分布,以狼毒为优势种的群落常在一些退化严重的草原上形成出现[6]。环境因子对高寒草甸不同坡度上狼毒盖度及狼毒群落盖度有着显著的影响,本研究结果显示,狼毒主要分布在海拔2900~3050 m、干燥阳坡15°~35°的高寒、亚高寒草甸或灌丛地带。其中,在土壤较厚、水分条件适中的地带狼毒长势较为旺盛,而在土壤湿度较大或较小的地带生长稀少。研究发现土壤水分对狼毒的生长分布有一定程度的影响,在一定区域内,当地表土壤水分含量越高时,狼毒长势越差[33]。本研究结果与其不完全一致,主要原因可能是本研究区位于青藏高原东北部边缘,受地形、地貌、光照等微生境及微气候条件的影响,导致研究结果具有一定的差异性。另外,坡度作为主要的地形因子能对草地土壤厚度、土壤水分含量及土壤侵蚀程度产生重要影响,从而使土壤养分表现出在不同坡度上的分异性[23],这也是研究区狼毒在不同坡度呈现分布差异的主要原因之一。
植物功能群是草原生态系统响应环境变化的基本单元,能够反映植物群落结构及生态系统功能机制在环境影响下的动态变化[34]。通过观察发现,调查样方中物种以莎草科植物和多年生杂类草居多,其中狼毒数量占总物种数量的17.7%,超过禾本科植物数量的10.9%,表明狼毒在亚高寒草甸中的比重在增多,这是由于退化草地在过度放牧后,原生植被群落结构和成分都发生了相应改变。在牛、羊等家畜啃食践踏及人为因素干扰影响下,禾本科和莎草科牧草生长潜力受到抑制,数量和质量不断下降,从优势种渐变为伴生种。而毒杂草则从伴生种不断蔓延扩散转变为优势种,这与王宏生等[2]的研究结果一致。狼毒型退化草地是青藏高原毒杂草型退化草地的主要类型之一,遗传多样性和繁殖优势使得狼毒成为退化草地上一种竞争力极强的毒草[9]。狼毒因具有毒性导致牲畜不采食,且以其具有发达的根系能够适应退化草地的土壤水分和养分环境[35-36]。同时,狼毒具有多实性能,可产生大量种子,对禾本科等类牧草具有较强的化感作用,能抑制它们的生长[37],而狼毒种群在原有分布区域内,其密度范围逐渐扩大并转变为优势种群,从而造成天然草地群落结构改变和植被退化。本研究在对不同坡度狼毒种群调查研究中发现,Ⅲ和Ⅳ坡度狼毒以建群种存在且重要值最高,其他植物重要值所占比重相对较小,且随着人类活动和牲畜的影响其范围在不断扩大,由原先的稀疏型分布逐渐演变为密集型分布。随着狼毒覆盖度的增加,群落中原有的优良牧草优势度逐渐下降,而狼毒等毒杂草优势度逐渐增加,成为群落的优势种[38],这主要是因为相对于其他物种,狼毒更具有生长和繁殖的竞争优势。研究表明退化程度重的草地群落中牧草的地上生物量和重要值随着狼毒分盖度的增加会出现明显下降[39],本研究结果与其一致。植物分布格局与地形因素关系密切,坡度是影响植物生境要素(光照、水文等)差异和植物分布的主要地形因子之一,在群落植被演化过程中具有一定地位[40]。坡度可通过改变生境局部区域的热量、水分、光照强度及土壤养分等非生物因子,对植物功能群产生直接或间接影响[23]。本研究调查发现狼毒群落植物功能群的分布与坡度有一定的关系,随着坡度的增大狼毒的分布逐渐减少,而莎草科植物和禾本科植物则逐渐增多,当坡度达到一定程度时植物的株高明显变低并出现匍匐状植物和裸露地块,植物生长表现出对不同坡度微生境的生态适应性,这主要是不同坡度上其土壤养分、水热条件等存在差异,使得研究区域出现了小范围的生境异质性,从而造成狼毒群落植物多样性在不同坡度的分异。
在植物与环境的相互关系中,随着环境的变化,植物能够调节环境适应策略以顺应环境变化带来的影响,形成适应环境变化的植物功能性状[41]。本研究发现,随着样地坡度的变化,植物的功能性状也发生相应的变化,主要表现在有些植物生长比较低矮,如蒲公英;有些植物具有内卷的叶片,如异针茅;有些植物具有灰白色绒毛,如冷蒿、猪毛蒿等;有些植物呈垫状,如薄雪火绒草。这些植物形态特征的不同都是物种适应环境生存的表现。本研究与已有研究结果基本一致,即随着生境条件的改变,土壤温湿度、养分等条件都可能发生变化,植物的功能性状也会发生适应性的变化,生境异质性是影响植物功能性状发生改变的主要环境因子[41-42]。亚高寒草甸植物功能性状的改变主要是为了适应研究区干旱、寒冷、多风的高寒环境及不同坡度土壤养分和水热条件差异造成的微生境异质性。
生物多样性是维系生态系统信息传递和流动物质循环的重要环节,包括遗传多样性、物种多样性、生态系统多样性和景观多样性[43],其中物种多样性是生物多样性的核心,也是生物多样性最主要的结构和功能单位[44]。物种多样性能够反映草地生态系统的基本状况,其核心是物种的数量变化以及生物学的多样性指数[45],物种多样性高的区域其群落结构也复杂[46]。物种多样性指数能够表征群落的结构类型、稳定程度以及反映其和生境之间的差异和联系,其数值大小受到了群落中植物物种丰富度和均匀度共同的影响。本研究中,由于受微地貌、光照、土壤养分、水热条件等因素影响,不同坡度植物群落物种组成存在显著差异。调查样地植物群落丰富度指数在不同坡度上变化幅度较大,不同坡度物种丰富度大小为Ⅳ>Ⅰ > Ⅱ>Ⅲ> Ⅴ,Shannon-Wiener多样性指数和Pielou均匀度指数变化为Ⅳ>Ⅴ>Ⅰ >Ⅱ> Ⅲ。结果显示,Ⅳ坡度上物种丰富度最高,植被分布最为均匀,究其原因是该坡度上生境相对复杂,水热、土壤养分等条件受微气候和环境等因素的影响变化多样。研究结果表明,由于不同坡度生境存在差异,其物种组成也明显不同,生境结构复杂的群落其多样性指数和物种丰富度指数高,Shannon-Wiener多样性指数和Pielou均匀度指数结果均显示出生境结构复杂的群落其物种多样性也相对丰富。
4 结论
在生态地理分布研究中发现,研究区狼毒主要分布在海拔2900~3050 m的高寒、亚高寒草甸地区。其中,在土壤养分良好、水分状况适中的15°~35°的干燥阳坡地带狼毒分布较多。在调查样地中,共记录到植物约50种。
从植物功能性状来看,为适应研究区干旱、寒冷、多风的高寒环境以及海拔和坡度的变化对生长环境的影响,不同植物物种在形态结构上具有明显不同的特征,具体表现在有些物种生长比较低矮,有些物种具有内卷的叶片,有些植物具有灰白色绒毛,有些植物呈垫状,这些都是物种适应环境生存的表现。
从植物功能群重要值变化看,莎草科植物重要值的分布为Ⅱ> Ⅳ>Ⅰ> Ⅴ>Ⅲ,最高值在坡度Ⅱ,最低值在坡度Ⅲ;多年生根茎禾草重要值的分布为Ⅴ> Ⅳ> Ⅲ>Ⅰ> Ⅱ,最高值在坡度Ⅲ,最低值在坡度Ⅱ;多年生丛生禾草重要值分布为Ⅴ>Ⅳ>Ⅰ> Ⅲ > Ⅱ,最高值在坡度Ⅴ,最低值在坡度Ⅱ;灌木与半灌木重要值分布为Ⅳ> Ⅴ>Ⅲ> Ⅱ>Ⅰ,最高值在坡度Ⅳ,最低值在坡度Ⅱ;多年生杂类草重要值分布为Ⅲ> Ⅱ> Ⅳ> Ⅴ>Ⅰ,最高值在坡度Ⅲ,最低值在坡度Ⅰ;一、二年生植物重要值分布为Ⅴ>Ⅳ> Ⅲ> Ⅱ > Ⅰ,最高值在坡度Ⅴ,最低值在坡度Ⅰ。研究区大多数植物功能群在坡度Ⅳ和Ⅴ上重要值最高,在坡度Ⅰ和Ⅱ上重要值最低,不同植物功能群表现出对不同坡度的选择差异性。
调查样地不同坡度上其微生境结构和植被群落物种组成存在差异,植物群落丰富度指数在不同坡度上变化幅度较大,生境结构复杂的群落其多样性指数和物种丰富度指数也高,Shannon-Wiener多样性指数和Pielou均匀度指数结果也显示出结构复杂的生境群落,其物种多样性相对丰富,坡度对亚高寒草甸狼毒群落植物多样性分布格局有一定的影响。
