水稻种子蛋白质的组成和积累形态对稻米品质的影响
2021-06-15刘慧芳吴孚桂聂佳俊韦云飞马启林
刘慧芳 吴孚桂 聂佳俊 韦云飞 马启林

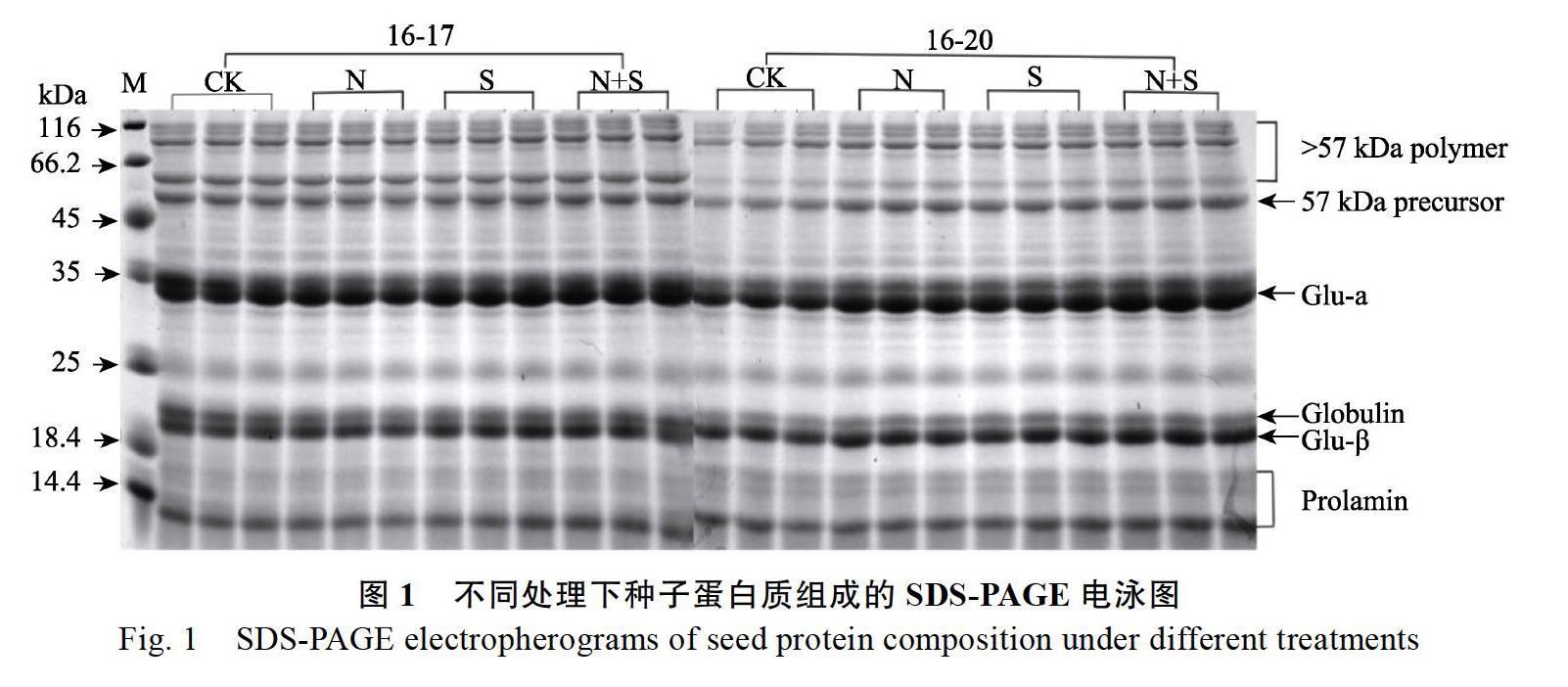
摘 要:設置水培试验对‘16-17(谷蛋白前体聚合物明显,含量高)和‘16-20(谷蛋白前体聚合物不明显,含量低)分别进行CK(氮2800 μmol/L,硫80 μmol/L)、N(氮3640 μmol/L,硫80 μmol/L)、S(氮2800 μmol/L,硫260 μmol/L)、N+S(氮3640 μmol/L,硫260 μmol/L)处理,以此改变稻米品质和稻米贮藏蛋白质的组成与含量,从而对稻米蛋白质各组分及相关品质性状进行分析。结果表明:(1)在N、S和N+S的不同处理下均增加了粗蛋白的含量,使‘16-20的稻米粗蛋白百分含量分别增加2.35%、2.67%和2.73%;(2)在2个品种的各组分蛋白含量变化中,‘16-17的醇溶蛋白和谷蛋白的百分含量在N、S和N+S下较对照分别增加了9.75%、4.21%和4.67%,3.92%、7.38%和12.92%。‘16-20的谷蛋白百分含量在S和N+S处理下增加了17.72%和29.38%,醇溶蛋白百分含量在N+S下增加了3.73%,而球蛋白百分含量在N、S和N+S三个处理下降低了2.86%、3.78%和3.69%,清蛋白在N、S处理下显著降低;(3)谷蛋白的2个亚基的相对含量和57 kDa谷蛋白前体及57 kDa以上谷蛋白聚合体的积累在‘16-20品种中明显增加。(4)稻米品质中,N、S配施对2个参试品种的碾磨品质性状的影响不明显,但有利于2个品种稻米外观品质和食味品质的改善,并使‘16-20的营养品质显著增加,另外,‘16-20的千粒重在S和N+S处理下显著增加。故不同N、S配施处理下,不同品种水稻贮藏蛋白质的组成和积累形态差异很大,并改变了含巯基蛋白质组分的积累量和积累形态。尤其是在富S条件下,蛋白质积累过程中形成较多的巯基及二硫键,并通过二硫键来改变含巯基的蛋白质亚基之间的聚合程度,形成不同分子量的谷蛋白聚合体,使蛋白质的积累形态发生明显改变,进而使稻米相关品质性状发生改变。
关键词:水稻;种子蛋白;蛋白组分;稻米品质
中图分类号:S511 文献标识码:A
Effects of Rice Seed Protein Composition and Accumulation Morphology on Rice Quality
LIU Huifang, WU Fugui, NIE Jiajun, WEI Yunfei, MA Qilin*
College of Tropical Crops, Hainan University, Haikou, Hainan 570228, China
Abstract: A hydroponic test is set for the processing of CK (nitrogen 2800 μmol/L, sulfur 80 μmol/L), N (nitrogen 3640 μmol/L, sulfur 80 μmol/L), S (nitrogen 2800 μmol/L, sulfur 260 μmol/L) and N+S (nitrogen 3640 μmol/L, sulfur 260 μmol/L) for the ‘16-17 (with high content of the precursor polymer of glutelin) and ‘16-20 (with low content of the precursor polymer of glutelin), by changing the rice quality and the composition and content of rice storage protein, in order to analyze the rice protein components and related quality traits. The crude protein content increased under different processing of N, S and N+S, that of ‘16-20 significantly increased by 2.35%, 2.67% and 2.73%. The content of prolamin and glutelin of ‘16-17 increased by 9.75%, 4.21% and 4.67%, 3.92%, 7.38% and 12.92%, respectively for the two varieties. The glutelin content of ‘16-20 significantly increased by 17.72% and 29.38% under the processing of S, the prolamin content increased by 3.73% under the processing of N+S,the percentage of globulin was reduced by 2.86%, 3.78% and 3.69% under the processing of N, S and N+S., and the albumin significantly reduced under the processing of N and S. The relative content of the two subunits of glutelin, 57 kDa glutelin precursor and glutelin polymer obviously increased in ‘16-20. With regard to rice quality, the combined application of N and S didnt generate obvious impact on the grinding quality of the two varieties, but was conductive to improving the appearance quality and taste quality of the two varieties, and significantly increased the nutritional quality of ‘16-20. Besides, the thousand seed weight of ‘16-20 significantly increased under the processing of S and N+S. Hence, under the combined application of N and S, the protein stored in different varieties of rice has great difference in the component and accumulation form, and changes the accumulation amount and accumulation form of the compounds containing sulfydryl proteins. Especially in the condition of abundant S, the protein forms much sulfydryl and disulfide bond in the accumulation process and changes the polymerization contents among protein subunit containing the sulfydryl through disulfide bond to form glutelin polymer with different molecular weight. As a result, the accumulation form of the protein has obvious change, further changing the relevant quality of rice.
Keywords: rice; seed protein; protein component; rice quality
DOI: 10.3969/j.issn.1000-2561.2021.04.030
水稻(Oryza sativa L.)是全球一半以上人口的主要粮食作物,也是植物蛋白的主要来源,它为全世界超过30亿的人提供了25%的能量[1-3]。而随着生活质量的逐步改善,人们对优质大米的需求今后也会强劲增长。水稻种子的主要营养成分是淀粉、储藏蛋白质、氨基酸、脂质等,其中大部分是淀粉和蛋白质,它们的重量合计占谷物干物质重量的90%以上[4-5]。大米中含有直链和支链2种淀粉,并且2种淀粉的形态结构和所占比例对大米的许多品质特性有重要影响[6]。除淀粉之外,水稻种子中的蛋白質和氨基酸含量以及它们的相对平衡是决定水稻营养价值的最重要因素[3, 7]。大米中的储藏蛋白质含量和组成对其蒸煮食味品质、营养价值、大米的外观和加工品质等都有重要的影响[8]。
水稻种子的蛋白质含量属于典型的数量性状,遗传变异丰富,同时品种间的遗传基础差异也非常大,并且容易受不同环境因素和栽培技术的影响[9]。不同施肥处理对水稻产量、食味品质及蛋白质组分的影响不同,其中以氮素的施用对其影响最大[10],因为氮素是蛋白质成分中的重要元素之一。另外,硫也是构成蛋白质的主要成分之一,提高硫素水平可以提高巯基含量,使含有-SH 氨基酸中半胱氨酸和甲硫氨酸通过-S-S-形成高分子量的聚蛋白,进而使其蛋白的结构和功能发生相应变化。并且,由于氮和硫的密切关系,在氮供应充足的条件下,硫肥的施用往往对产量都有明显的增产效应,Luo等[11]的研究发现,在仅仅施用硫肥的情况下稻米的相关品质不会发生显著变化,硫肥对稻米品质的影响只有在施加一定量氮肥的前提下才会发生明显的改变。
关于水稻蛋白质各组分与稻米品质的关系方面的研究较少,现有的研究主要集中在氮、硫对水稻产量和粗蛋白含量的影响,以及氮、硫对水稻稻米品质的影响,而对水稻中各蛋白的代谢与积累形态对稻米品质影响的研究尚未见详细报道。另外,蛋白的其他组分对稻米品质的影响如何尚不清楚。本研究在水稻生产过程中,通过氮、硫配施,改变水稻种子蛋白的含量、组成和积累形态,进而分析对稻米品质有何影响,旨在研究水稻种子蛋白积累形态对稻米品质的影响,从而为提高水稻种子蛋白含量并改善稻米品质提供参考。
1 材料与方法
1.1 材料
以蛋白质组分和含量不同的2个水稻品种‘16-17(谷蛋白前体聚合物明显,含量高),‘16-20(谷蛋白前体聚合物含量低)作为试验材料。
1.2 方法
1.2.1 种植方法 将试验材料于2019年7月9日播种,湿润育秧;7月21日移栽到水田中,长至开始出现分蘖;8月25日移至水培溶液中,采用温室塑料钵栽培,塑料钵的规格为口径20 cm,高30 cm,每钵装2 kg干净的细沙,然后放入装满营养液的培养池中,置于温室大棚中培养,直至成熟。营养液(国际水稻水培营养液)分为4个处理,分别为CK(N含量为2800 μmol/L,S含量为80 μmol/L),N(N含量为3640 μmol/L,S含量为80 μmol/L),S(N含量为2800 μmol/L,S含量为260 μmol/L),N+S(N含量为3640 μmol/L,S含量为260 μmol/L)。其中,S含量80 μmol/L是水稻生长所需S元素的临界值含量,S含量260 μmol/L是水稻生长所需S元素的相对最适含量;N含量2800 μmol/L是国际水稻水培营养液正常含量,N含量3640 μmol/L是在S含量最适的情况下,根据N∶S=14∶1计算所得。成熟后收获干燥的种子,烘干至恒重后,取稻穗中部颗粒饱满、形状大小一致的籽粒,去壳,研磨成细粉,分别进行粗蛋白和蛋白质各组分的提取与测定。以每5粒干燥籽粒为1组,取3组平均值为籽粒单粒重,粗蛋白及蛋白质各组分的提取与测定均为3次重复,以占籽粒干重的百分率为单位[12]。
1.2.2 粗蛋白提取和测定 参照Takemoto等[13]的方法略有改动。称量研磨后的水稻米粉,放入2 mL的离心管中,按照20 mg/ml(W/V)的量加入SDS-Urea蛋白提取液[三羟甲基氨基甲烷(Tris-base)0.37815 g,HCl调节pH到6.8;尿素6 g,十二烷基硫酸钠(SDS)0.25 g,2-巯基乙醇 1.25 mL,加蒸馏水定容到100mL]。振荡混匀后放置MX-RL-标准型旋转混匀仪上旋转过夜,于14000 r/min、4 ℃进行离心15 min,取上清液用Bradford试剂盒, 测定595 nm处的吸光值,计算粗蛋白含量。
1.2.3 蛋白质各组分的提取和测定 参照Takemoto等[13]的方法,有改进。水稻米粉用丙酮脱脂后依次用H2O、1.0 mol/L磷酸缓冲液、60%异丙醇、1%乳酸溶液连续提取清蛋白、球蛋白、醇溶蛋白及谷蛋白4种不同组分。蛋白质含量测定同粗蛋白的测定方法一致。
1.2.4 SDS-PAGE聚丙烯酰胺凝胶电泳 采用14%的分离胶,5%的浓缩胶。取10~20 ?L蛋白变性液上样,进行100 V、3 h左右的SDS-PAGE电泳,考马斯亮蓝R-250染色、脱色,成像观察。
1.2.5 稻米品质相关测定 参照国家标准GB/T 1354—2018《大米》和NY/T 593—2013《食用稻品种品质》测定所取样品的精米长、精米宽、长宽比,垩白米率、糙米率、精米率、整精米率、直链淀粉(索莱宝直链淀粉试剂盒)和胶稠度等稻米品质的相关指标。
1.2.6 二硫键和巯基含量的测定 游离巯基(SHF)、总巯基(SHT)和二硫键(-S-S-)的含量均采用Beveridge等[14]的方法测定,并稍作改动。称取60 mg磨碎的米粉溶于10 mL Tris-Gly-8M Urea缓冲液,混匀后10000 ×g离心10 min,取上清液测定游离巯基(SHF)和总巯基(SHT)含量。
游离巯基(SHF)含量:取2 mL上清液于新的离心管,向其加入80 μL Ellmans试剂,然后立即混匀,5 min后在412 nm下测吸光值。总巯基(SHT)含量:取2 mL上清液于新的离心管中,加入0.2% β-巯基乙醇,摇匀静置2 h,再加入4mL 12%的三氯乙酸(TCA),摇匀后静置1 h沉淀蛋白质,10000 ×g离心10 min,用12%的TCA溶液洗涤沉淀4次,最后将沉淀溶于3 mL Tris-Gly缓冲液中,取2 mL测吸光值(A412)。计算公式如下:
2 結果与分析
2.1 不同营养液对不同水稻品种蛋白组分含量的影响
表1的结果显示,对不同品种而言,不同处理下蛋白组分的变化不同。与对照相比,‘16-17水稻品种在N、S和N+S的处理下,清蛋白、球蛋白含量均下降,但变化不显著,而醇溶蛋白和谷蛋白的含量则显著增加,醇溶蛋白在N处理下由14.22%提高至23.97%。谷蛋白的百分含量在N+S处理下显著增加,增加了12.97%。N、S处理使醇溶蛋白和谷蛋白含量显著增加,其他蛋白相对含量降低。对‘16-20水稻品种在不同的处理下,清蛋白含量在N、S处理下显著降低,百分含量降低了2.52%、2.56%。球蛋白含量在N、S、N+S处理显著降低,百分含量分别下降2.86%、3.78%和3.69%。醇溶蛋白的百分含量在N+S处理下显著增加,百分含量增加了3.73%。谷蛋白的百分含量在S和N+S处理下显著增加,分别增加了17.72%和29.38%。所以在N、S、N+S处理下显著降低清蛋白和球蛋白含量,显著增加谷蛋白含量。
2.2 不同营养液处理改变蛋白的积累形态
表1的结果显示,不同处理后,2个品种的谷蛋白的相对含量均有所增加。‘16-17经不同处理后,醇溶蛋白相对含量显著增加,清蛋白和球蛋白变化不显著。而‘16-20品种的醇溶蛋白在N+S处理下显著增加,清蛋白、球蛋白的含量均降低。由此可见,对不同水稻品种而言,在不同处理下各组分蛋白的组成变化有很大差异。根据
SDS-PAGE的结果(图1)显示,57 kDa谷蛋白前体含量高的‘16-17,各组分蛋白在施加不同的营养液处理时,除了清蛋白和醇溶蛋白有所变化,球蛋白和谷蛋白组分变化没有明显差异;而57 kDa谷蛋白前体含量低的‘16-20品种,蛋白含量有明显的差异,尤其是谷蛋白的含量明显增加,并且增加了57 kDa及以上多肽的积累。这可能主要是因为谷蛋白的前体物质,即前原蛋白及大分子谷蛋白聚合体的形成具有密切的关系,由于聚合体的形成而造成水稻种子谷蛋白的酸性和碱性2个亚基的含量发生相应的变化,根据结果显示谷蛋白的α-亚基变化最明显。并且可以推测,谷蛋白的α-亚基的多肽链的组成也极有可能发生了改变。这说明N、S处理后的水稻蛋白,不仅谷蛋白的相对含量发生变化,而且谷蛋白的积累形态、谷蛋白的前体物质前原蛋白及大分子谷蛋白聚合体也明显地发生了改变。
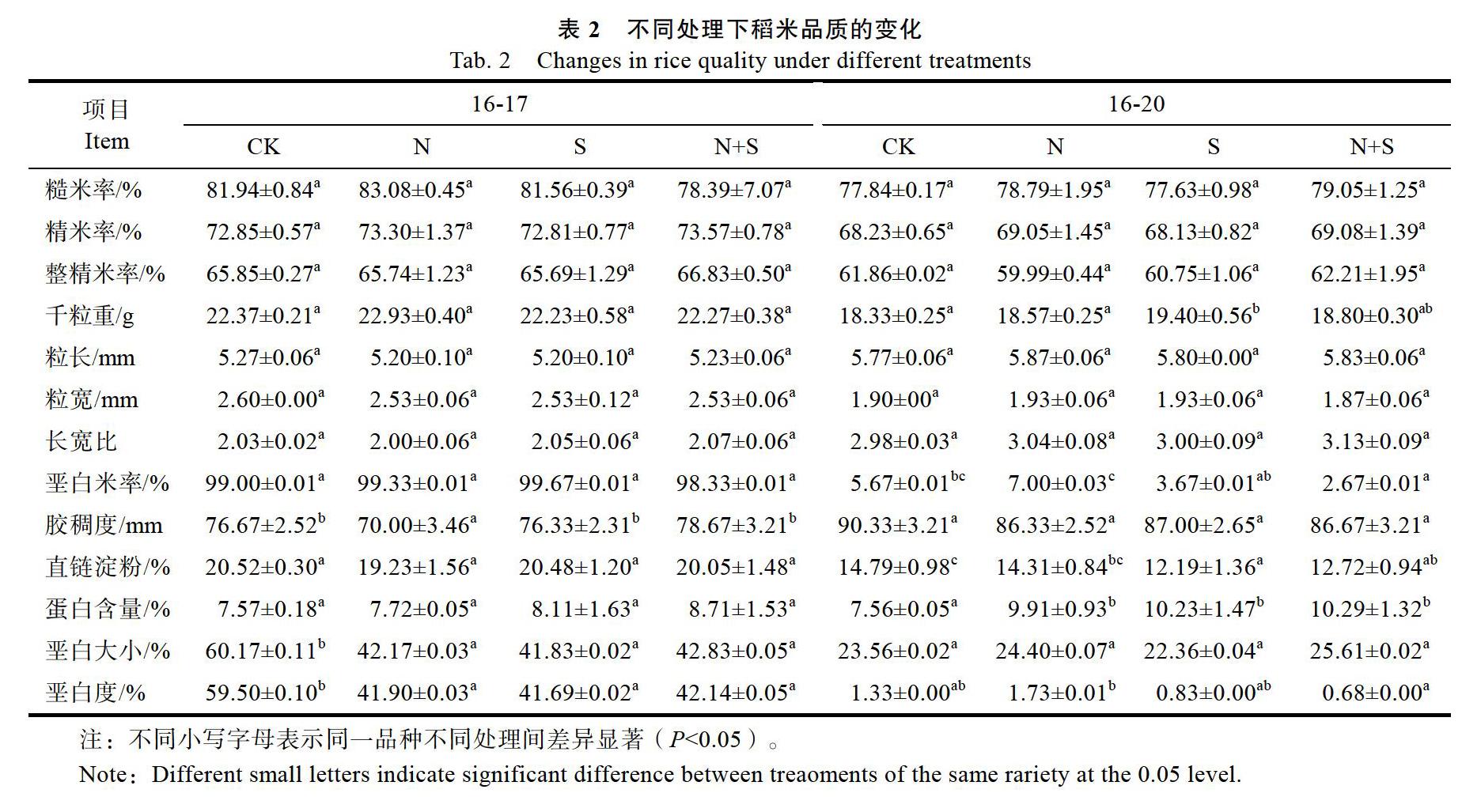
2.3 不同营养液处理对稻米品质的影响
根据表2结果显示,在碾磨品质方面,对于‘16-17和‘16-20,不同处理条件下,糙米率、精米率和整精米率均无显著性差异。在外观品质方面,‘16-17和‘16-20的粒长、粒宽和长宽比差异不显著;另外,不同处理下‘16-17的垩白米率差异不显著,但是其垩白大小百分比在N处理下显著降低18%,在S处理下显著降低18.34%,在N+S处理下显著降低17.34%,垩白度也显著降低。‘16-20在施加N处理下垩白米率显著增加,在S处理下百分含量降低2%但是变化不显著,在N+S处理下百分含量显著降低3%,而垩白大小没有显著差异,垩白度与CK相比没有显著差异,但是N和N+S之间存在差异,N+S处理下的垩白度显著低于N处理下的垩白度。在蒸煮品质方面,不同处理下,‘16-17的直链淀粉含量差异不显著,‘16-20的直链淀粉含量为CK>N>N+S>S,其中在S和N+S处理下直链淀粉含量显著降低,与对照相差分别为2.6%、2.07%。胶稠度方面,‘16-20稻米的胶稠度在不同处理下变化不显著,但‘16-17的胶稠度在施加N处理下为70 mm,显著低于其他处理。在营养品质方面,‘16-17的蛋白含量CK 2.4 不同营养液处理下巯基、二硫键的变化 表3的结果显示,对于‘16-17,总巯基含量为CK 3 讨论 3.1 不同N、S配施处理下种子蛋白质变化的差异 相关研究[13]发现,谷蛋白和醇溶蛋白作为水稻贮藏蛋白的主要成分,分别积累在2种不同的蛋白体PB-II和PB-I中。为了达到水稻稻米蛋白与不同品质性状间的完美结合,需要我们精准掌握各个水稻品种的不同蛋白组分的调节,所以在这方面研究仍有无限可能[15]。一般认为,通过外界施加氮肥、硫肥会明显提高水稻籽粒中蛋白质及其各组分的相对含量[16-18]。从本研究结果可以看出,施加N、S可以增加水稻蛋白质含量,但不同品种之间又存在明显差异,‘16-17的种子总蛋白含量增加不显著,‘16-20的种子总蛋白含量则显著增加。其次,种子蛋白质各组分随着N、S的施用在不同品种之间也存在明显差异,‘16-17在N、S处理下清蛋白、球蛋白变化不显著,而与之相反的醇溶蛋白和谷蛋白含量显著增加。这与Ning[19]等的研究结果相同,即外界施加氮素等对醇溶蛋白和谷蛋白含量变化的影响更大。对‘16-20而言,在N、S、N+S的处理下,清蛋白,球蛋白的含量显著降低,醇溶蛋白没有的研究结果有所不同,可能是因为‘16-20水稻显著变化,而谷蛋白的含量显著增加。这与前人种子的总蛋白和谷蛋白含量增加,而含硫氨基酸主要作用于谷蛋白,故醇溶蛋白的变化不明显,而清蛋白和球蛋白含量显著降低,这方面的变化需要进一步深入研究。从电泳图和蛋白组分测定的结果可见,‘16-20的总蛋白和各组分蛋白更容易受外界施加的N、S的影响。 3.2 不同N、S配施處理下稻米品质的变化 本研究通过水培方法探讨了不同N、S配施条件下稻米品质的变化,结果与土壤栽培条件下施加氮、硫肥对稻米品质的影响,既有表现一致的一面,又有表现不一致的一面。李玉影[18]在土壤栽培条件下的研究指出,增施硫肥具有增加稻米蛋白质含量、降低直链淀粉含量的作用;王爽等[17]提出外界增加氮肥施用量可以提高水稻产量,同时增加稻米的碾磨品质和蛋白质含量,降低胶稠度和直链淀粉含量,这与本文的水培试验结果一致。不论是单独施加N或S的处理,还是N+S配施处理,2个试验品种的种子蛋白质含量都呈增加的趋势,特别是‘16-20的种子蛋白质含量更是显著性地增加。而碾磨品质在不同N、S配施条件无显著性变化。外观品质方面,垩白米率、垩白大小和垩白度表现出明显差异,S或N+S的施用条件下,2个品种的垩白米率和垩白度都有减少,有利于外观品质的改善,这与陈能等[20]蛋白质含量对稻米的垩白度、垩白米率等质量指标产生不良影响的结果相反,可能是因为该研究的2个水稻品种不属于高蛋白品种,故其外观品质较高蛋白品种更优一些。蒸煮品质方面,在S处理下‘16-20品种的直链淀粉含量显著降低,这与陈小倩等[21]的研究结果一致,一般认为在一定范围内直链淀粉含量降低则食味值增加。本研究的结果初步显示,单独S处理,以及N+S配施处理,均有利于改善‘16-20的稻米营养品质和外观品质,对于其他品质的影响还需进一步的研究。 3.3 含巯基的蛋白与稻米品质的相关性 水稻种子蛋白组分中,谷蛋白和醇溶蛋白都富含半胱氨酸和蛋氨酸,这2个氨基酸都含有自由巯基,这些巯基可以通过分子内或分子外的二硫键相互结合而形成较大的蛋白质聚合体[22],这些蛋白质聚合体进一步与淀粉粒结合形成蛋白质- 淀粉复合体等网络结构,可能成为影响米饭硬度和黏性等食味品质的主要作用之一[23-24]。本研究结果显示,施加S处理下游离巯基、总巯基和二硫键含量都有所增加,尤其是二硫键含量显著增加的‘16-20的谷蛋白含量也显著性增加。水稻种子中的谷蛋白和醇溶蛋白的结构和特性主要是由二硫键决定的, 并对2种蛋白的结构性和稳定性起着至关重要的作用[25]。硫对水稻种子蛋白质含量的影响主要体现在,施加硫元素能够增加水稻蛋白质含量,并对蛋白质的积累形态产生影响,这种影响主要集中在含巯基的谷蛋白和醇溶蛋白的质和量的变化上。在施硫处理下,变化最明显的是谷蛋白亚基、57 kDa谷蛋白前体和大分子蛋白聚合物的含量和形态,从SDS- PAGE电泳图谱上看,施S处理下这几种形态的蛋白质含量显著增加,尤其是‘16-20的57 kDa的大分子的形成和含量的增加很明显,100 kDa左右的蛋白条带变化较明显,并且醇溶蛋白的变化也有一定差异。硫处理下,2个品种的清蛋白和球蛋白含量降低、谷蛋白含量增加,而醇溶蛋白含量因品种的不同而有不同的变化,这一差异可能影响水稻种子蛋白组分的变化,而水稻蛋白组分间比例的变化也可影响蛋白质最终的结构性质、功能性等的改变,从而导致稻米品质的改变。 此外,根据稻米品质相关分析显示,变化最明显的为淀粉,并且有关研究[26-27]表明蛋白质与淀粉、淀粉与脂质之间的相互作用、结合构成的某种网络结构对大米的蒸煮、糊化和胶稠度等都有影响,对大米的性质造成改变。正是由于这种含巯基的蛋白质结构对蛋白质、淀粉、脂质作用而复合形成的结构,直接或间接地影响稻米的蒸煮食味品质的质地和大米的营养品质。 4 结论 研究结果显示,氮、硫的施用可以显著改变水稻种子蛋白质含量,使蛋白质的4种组分的含量和形态也发生明显变化。硫的施用可以显著增加总巯基含量和二硫键含量,导致某些富含含硫氨基酸的蛋白质组分合成增加,并通过二硫键结合,使水稻种子蛋白质的结构和积累形态发生改变。同时氮、硫配施也改变了稻米的品质性状,同时其变化程度与品种有一定的关系,结果显示品种‘16-17和‘16-20间的差异也很大。另外,在硫的施用下而引起的水稻种子蛋白质成分和积累形态的改变与稻米的品质性状有密切关系。 参考文献 [1]Kusano M, Yang Z, Okazaki Y, et al. Using metabolomic approaches to explore chemical diversity in rice[J]. Molecular Plant, 2015, 8(1): 58-67. [2]Tian Z, Qian Q, Liu Q, et al. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(51): 21760-21765. [3]Peng B, Kong H L, Li Y B, et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice[J]. Nature Communications, 2014, 5: 4847. [4]He Y, Wang S, Ding Y. Identification of novel glutelin subunits and a comparison of glutelin composition between japonica and indica rice (Oryza sativa L.)[J]. Journal of Cereal Science, 2013, 57(3): 362-371. [5]彭 波, 庞瑞华, 孙艳芳, 等. 水稻种子淀粉合成及其调控研究进展[J]. 江西农业学报, 2016, 28(6): 15-21. [6]Hyun D Y, Lee G A, Kang M J, et al. Development of low temperature germinability markers for evaluation of rice (Oryza sativa L.) germplasm[J]. Molecular Breeding, 2015, 35(4): 104. [7]Duan M, Sun S S. Profiling the expression of genes controlling rice grain quality[J]. Plant Molecular Biology, 2005, 59(1): 165-178. [8]曹珍珍, 张其芳, 韦克苏, 等. 水稻籽粒氮代谢几个关键酶对花后高温胁迫的响应及其与贮藏蛋白积累关系[J]. 作物学报, 2012, 38(1): 99-106.. [9]石 吕, 张新月, 孙惠艳, 等. 不同类型水稻品种稻米蛋白质含量与蒸煮食味品质的关系及后期氮肥的效应[J]. 中国水稻科学, 2019, 33(6): 541-552. [10]兰 艳, 黄 曌, 隋晓东, 等. 施氮量对低谷蛋白水稻产量及品质的影响[J]. 华南农业大学学报, 2019, 40(4): 8-15. [11]Luo C, Branlard G, Griffin W B, et al. The effect of Nitrogen and Sulphur fertilization and their interaction with genotypeon wheat glutenins and quality parameter[J]. Journal of Cereal Science, 2000, 31(2): 185-194. [12]马启林, 李阳生, 鄢圣之, 等. 高温及穗肥对水稻子粒蛋白质积累动态的影响[J]. 湖北农业科学, 2008, 47(11): 1228-1231. [13]Takemoto Y, Coughlan S J, Okita T W, et al. The rice mutant esp2 greatly accumulates the glutelin precursor and deletes the protein disulfide isomerase[J]. Plant Physiology, 2002, 128(4): 1212-1222. [14]Beveridge T, Toma S J, Nakai S. Determination of SH- and SS-groups in some food proteins using Ellman's reagen[J]. Journal of Food Science, 1974, 39: 49-51. [15]朱永波, 韩展誉, 程方民. 氮肥对水稻营养品质、外观品质和加工品质的影响[J]. 基层农技推广, 2019, 7(12): 26-29. [16]张 欣, 施利利, 刘晓宇, 等. 不同施肥处理对水稻产量、食味品质及蛋白质组分的影响[J]. 中国农学通报, 2010, 26(4): 104-108. [17]王 爽. 氮肥用量和插秧密度对寒地粳稻蛋白质及稻米品质的影响[D]. 哈尔滨: 东北农业大学, 2015. [18]李玉影. 水稻需硫特性及硫对水稻产质量的影响[J]. 土壤肥料, 1999(1): 24-26. [19]Ning H F, Liu Z H, Wang Q S, et al. Effect of nitrogen fertilizer application on grain phytic acid and protein concentrations in japonica rice and its variations with genotypes[J]. Journal of Cereal Science, 2009, 50(1): 49-55. [20]陈 能, 罗玉坤, 谢黎虹, 等. 我国水稻品种的蛋白质含量及与米质的相关性研究[J]. 作物学报, 2006, 32(8): 1193-1196. [21]陈小倩, 曹伟召, 左 军, 等. 不同施氮量对水稻南粳46产量及品质的影响[J]. 现代农业科技, 2020(2): 17-18, 23. [22]马启林, 邓用川, 李阳生. 谷蛋白的聚合化特性及其对稻米品质的影响[C]//中国作物学会. 中国作物学会50周年庆祝会暨2011年学术年会论文集. 成都: 中国作物学会, 2011: 121. [23]Chrastil J, Zarins Z M. Influence of storage on peptide subunit composition of rice oryzenin[J]. Journal of Agricultural and Food Chemistry, 1992, 40(5): 927-930. [24]谢新华, 李晓方, 肖 昕. 蛋白质中二硫键对稻米淀粉黏滞性的影响[J]. 核农学报, 2009, 23(1): 114-117. [25]李 府, 王振林, 李文阳, 等. 施硫对小麦子粒形成过程中蛋白质与巯基、二硫键的含量及加工品质的影响[J]. 植物营养与肥料学报, 2011, 17(2): 291-300. [26]Paliwal J, Thakur S, Erkinbaev C. Protein-starch interactions in cereal grains and pulses[J]. Encyclopedia of Food Chemistry, 2019: 446-452. [27]Saleh M I. Protein-starch matrix microstructure during rice flour pastes formation[J]. Journal of Cereal Science, 2017, 74: 183-186. 責任编辑:崔丽虹
