黄鳍棘鲷放流苗种的遗传质量评估
2021-06-12朱克诚刘宝锁伞利择郭华阳江世贵张殿昌
朱克诚,刘宝锁,伞利择,刘 波,张 楠,郭华阳,郭 梁,江世贵,张殿昌
(1.中国水产科学研究院南海水产研究所农业农村部南海渔业资源开发利用重点实验室,广东 广州 510300;2.南方海洋科学与工程广东省实验室,广东 广州 511458;3.中国水产科学院南海水产研究所热带水产养殖研究开发中心,海南 三亚 572018;4.广东省海洋生物种业工程技术研究中心,广东 广州 510300)
在水生生物增殖放流活动中,利用人工繁育技术获得的水生生物苗种质量参差不齐,在被投放到天然水域后,随即与本水域中的自然群体混合。许多研究表明,在遗传多样性指标方面,假定放流群体与本水域自然群体存在一定差异,那么该自然群体的遗传结构、物种多样性和生态系统结构就有可能遭受破坏[1-5]。因此,针对放流苗种的遗传质量评估显得尤为重要。“放流苗种遗传质量”被定义为自然种群与放流苗种群体遗传多样性指标的吻合水平,对该放流而言,遗传多样性各项指标一致性越高,说明该放流苗种的遗传质量越好[6]。
我国增殖放流的评价体系还不完善,放流苗种遗传质量监测评估技术尚未健全,放流存在一定的盲目性,处于“粗放”的状态[7-8]。苗种放流评估研究方面,胡鹏飞等[9]在免疫、消化和营养等方面针对放流的中国明对虾苗种开展相关研究。此外,付亚男等[6]采用比对分析放流苗种和自然种群遗传多样性差异水平。
黄鳍棘鲷(Acanthopagrus latus)主要分布于我国南方近岸海域,近年来,由于环境污染、病害频发、捕捞量过大等促使其自然种群衰退明显[10]。国内,有学者已经开展关于黄鳍棘鲷增殖放流效果的评估以及标记物的筛选分析[11-13]。朱克诚等[10]利用微卫星亲子鉴定技术手段筛选的微卫星标记能为黄鳍棘鲷增殖放流、种群选育和分子辅助家系管理提供基础信息。基于生态系统通道(Ecopath) 模型,刘岩等[14]评估珠江口黄鳍棘鲷的生态容纳量,提出有针对性地进行增殖放流活动。吴仁协等[15]开发的47 个高多态性的黄鳍棘鲷微卫星标记可以为鲷科鱼类的种群遗传学分析提供标记来源。此外,有学者还从生理学[16-17]、病害[18-20]、肌肉发育[21-22]、性别调控[23]等方面解析其遗传基础。但是,关于放流前的黄鳍棘鲷苗种遗传质量评估的技术和方法未见报道。本研究参照付亚男等[6]的检测方法,以黄鳍棘鲷为研究对象,从遗传多样性水平、遗传分歧程度和近交程度等方面,比较分析黄鳍棘鲷自然群体和放流苗种群体的遗传质量,并详细描述该苗种的放流遗传特征,为鱼类放流苗种的遗传质量评估提供一定的参考依据。
1 材料和方法
1.1 生物样品采集与基因组总DNA 提取
2017 年12 月于中国水产科学研究院南海水产研究所热带水产研究开发中心实验采集黄鳍棘鲷幼鱼393 尾,平均体长(2.3±0.2)cm,该批苗种用于广东省阳西县沙扒镇海域增殖放流,其亲本来源于广东省阳西县沙扒海域附近海域的野生个体,数量为112 尾,雄鱼74 尾和雌鱼38 尾,平均体长(21.2±1.8)cm,平均体质量(514±36)g。2017 年12 月于广东省阳西县沙扒镇天然海域收集黄鳍棘鲷野生群体样品共计36 尾。全鱼和鳍条样品分别来自于仔鱼和野生群体,随即于无水乙醇中保存备用。
用酚氯仿方法[24]提取黄鳍棘鲷放流苗种与自然群体的基因组DNA,提取的DNA 样品最终稀释成100 ng/μL,并利用分光光度计和琼脂糖凝胶电泳进行DNA 浓度和质量的检测,后续-20 ℃保存待用。
1.2 微卫星等位基因分型
从该实验室前期开发的微卫星标记中选取12个黄鳍棘鲷标记[10],分别对黄鳍棘鲷自然和苗种群体样品进行等位基因分型,各标记的基本信息见表1。5′ 端带有HEX、TET 和FAM 的荧光引物均由上海生工生物工程技术服务有限公司合成。本实验随机将12 个SSR 标记分成4 组,标记组合及其信息见前期研究基础[10]。

表1 12 个微卫星标记引物特征Table 1 Primer characteristics of 12 microsatellite markers
PCR 反应体系参考已发表的文献描述[10],PCR产物检测方法同上。随后对PCR 产物进行Short tandem repeats (STR) 测序分析(上海生工生物工程技术服务有限公司,遗传分析仪,ABI3730XL 全自动DNA 测序仪),采用毛细管电泳技术检测每个个体的基因型,GS-500 作为标记。
1.3 种群遗传多样性和遗传质量
使用Arlequin version 3.5[25]计算观察等位基因数(NA)、观测杂合度(HO)、期望杂合度(HE)、分析苗种与自然群体间的固定指数FST;概率测试(probabilitily test)来统计检验FST值的差异显著性。多态信息含量(PIC)由PIC_CALC 0.6 软件来分析计算。软件GeneMarker V1.5 读取等位基因大小。
利用软件Coancestry 1.0[26]以“triadic likelihood estimator”模型来计算自然与苗种群体中个体的近交系数(inbreeding coefficient,F);使用软件Fstat 2.93 以“连锁不平衡法”(linkage disequilibrium methord)算法检测各群体的FIS。
2 结果
2.1 放流苗种和自然群体遗传多样性水平
黄鳍棘鲷放流苗种与自然群体的遗传多样性指标见表2。在放流苗种与自然群体中,利用12 个SSR 标记分别检测到119 和97 个等位基因。上述SSR 标记在苗种群体中NA的分布范围为5~ 16(平均值=9.917);上述SSR 标记在自然群体中NA的分布范围为2~ 17(平均值=8.083)。

表2 黄鳍棘鲷放流苗种与自然群体遗传多样性指标汇总Table 2 Summary of genetic variability of Acanthopagrus latus fingerling and natural stocks
苗种群体中各标记的HO为0.321~ 0.848(均值0.646),HE为0.394~ 0.850(均值为0.694);自然群体中各标记的HO为0.306~ 0.944(均值0.674),HE为0.306~ 0.911(均值为0.707)。以上结果表明放流苗种和自然群体中的HE均大于HO,说明以上两群体中的杂合子基因型存在缺失。
苗种群体中各标记PIC 为0.315~ 0.827(均值0.654);自然群体中各标记PIC 为0.296~ 0.890(均值0.659)。
2.2 种群近交程度
放流苗种群体F均值为0.050 5,自然群体F均值为0.034 2。
放流苗种群体FIS值为0.051 8,自然群体FIS值为0.036 5。
2.3 放流苗种遗传质量分析
该研究利用遗传多样性水平、遗传分歧程度、近交程度3 个方面中的6 个遗传指标,全面比较分析了放流苗种和自然群体的遗传属性,进一步评价黄鳍棘鲷放流苗种的遗传质量。
首先,对放流苗种和自然群体在4 个种群遗传多样性水平指标(NA、PIC 均值、HO均值、HE均值)(表3)进行了对比分析。结果表明自然群体的PIC 均值、HO均值、HE指标值均大于放流苗种群体,两者之间的偏差率的范围为-0.76%(PIC 均值)至 -4.15%(HO均值)。此外,放流苗种群体的NA均值高于自然群体的,偏差率为22.68%。
其次,种群间遗传分化检测结果显示放流苗种和自然群体间的遗传分歧指标呈现出显著差异(P<0.001),分歧程度定义为“微弱”(FST=0.011 2)。这表明在等位基因频率上,放流苗种群体与自然种群之间存在一定的微弱差异,其遗传结构仍存在差异。如果在该自然海域对黄鳍棘鲷苗种大量放流,则会导致自然种群等位基因频率的改变,进一步影响其遗传结构。
最后,结果表明放流苗种群体的F均值与FIS值均大于自然群体,两者之间的偏差率分别是47.66%和41.92%,表明在种群近交程度上,苗种群体大于自然群体。如果在该自然海域对黄鳍棘鲷苗种大量放流,则会使自然种群近交程度增加,造成自然种群遗传多样性信息加速流失。
总而言之,基于上述种群遗传学指标,黄鳍棘鲷放流苗种大部分皆逊于自然群体,说明在遗传质量方面,该苗种在遗传质量方面不符合放流要求。基于各项指标的偏差率,近交系数最高(41.92%~47.66%),该放流苗种最突出的遗传质量缺陷,此外HE和HO偏差率较高(-4.15%和-1.84%),说明该苗种遗传多样性方面也存在一定质量问题(表3)。
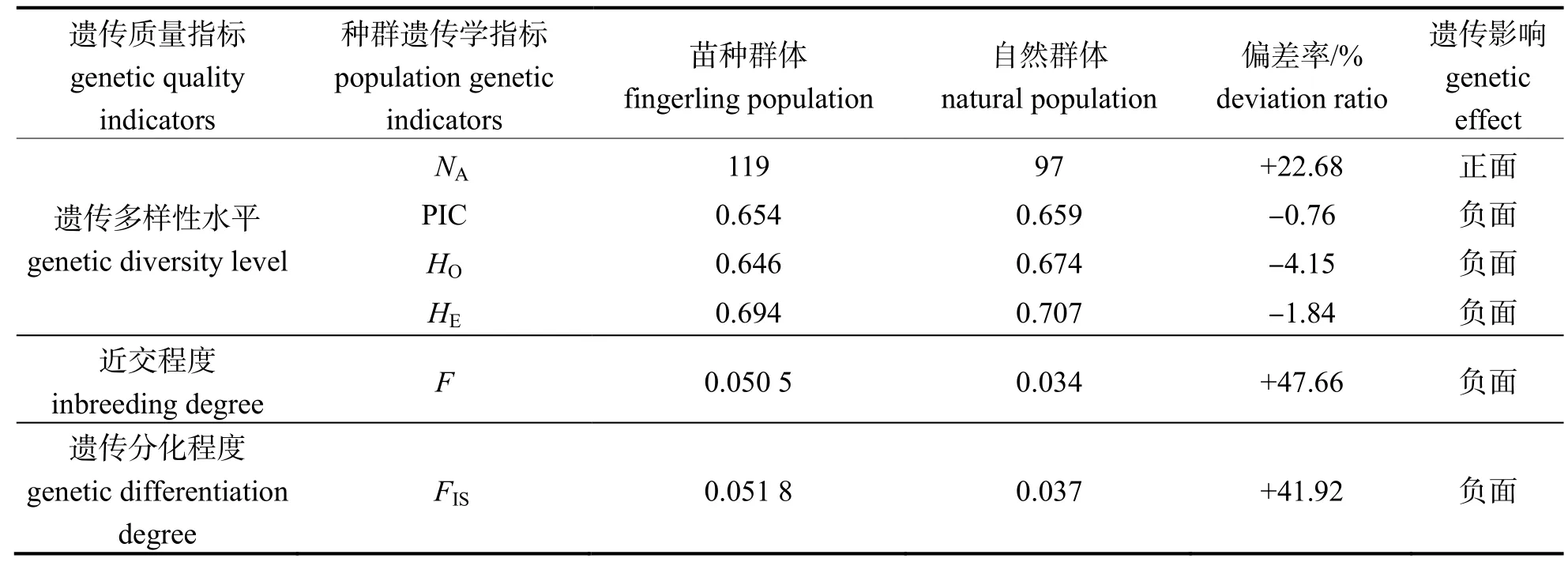
表3 黄鳍棘鲷放流苗种群体与自然群体遗传学属性比较Table 3 Comparisons of population genetic indicators between Acanthopagrus latus fingerling and natural stocks
3 讨论
3.1 放流苗种遗传质量评估的重要性
开展增殖放流、发展增殖渔业,是改善水域环境、优化渔业资源种群结构和质量、加强渔业资源恢复的一个重要途径。这对于近海渔业资源可持续利用、渔民增收、渔业增效、开拓渔业发展空间都具有十分重要意义。截至目前为止,已有许多学者指出自然种群遗传效应能受到水生生物增殖放流的影响[1-4]。其中,在山东半岛南部海域,中国对虾放流苗种对其野生资源起到了较好补充效应[27]。然而,也有Shan 等[5]在针对珠江河口黑鳍棘鲷的增殖放流苗种遗传质量评估研究中表明,与本水域自然种群相比较,黑鳍棘鲷的苗种群体遗传多样性水平较低,增殖放流后可能会影响自然群体的遗传结构水平。付亚男等[6]指出红鳍笛鲷(Lutjanus erythopterus)苗种的遗传多样性水平低于自然群体,不符合增殖放流中苗种遗传质量的需求。国外也有学者研究指出在近50 年内的70 项有关放流遗传结构方面影响的成果,其中证实自然种群遗传多样性程度优于放流苗种的有28 项,增殖放流对自然种群适应力起到了负面影响的有23 项[1]。
可见,评估放流苗种质量的优劣是当前迫切需要解决的任务。通过遗传评估信息,可检测自然与放流群体的遗传结构差异,可筛选掉遗传信息含量较少的放流苗种,以及进一步阻止含有较大差异遗传信息的放流苗种流入天然海域,从而破坏水生生物原有的遗传结构。
3.2 黄鳍棘鲷自然种群的遗传多样性
黄鳍棘鲷是中国南海区重要的经济海水鱼类之一,年养殖产量突破了6 万t。然而近年来,其自然种群严重衰退,遗传资源量显著下降[10,12,28],而生物种质资源的重要评价指标是生物遗传多样性水平[29]。但是目前关于黄鳍棘鲷自然群体遗传多样性的研究报道很少。朱克诚等[10]选用了12 对微卫星标记对广东省阳西县海域黄鳍棘鲷自然群体进行了遗传结构指标分析,结果表明利用分离出来的标记均具有丰富多态性。针对福建和珠江口2 个自然黄鳍棘鲷群体,杨慧荣等[28]利用RAPD 技术对其进行遗传变异分析,结果表明珠江口和福建群体内的遗传变异水平都较高,但珠江口群体的遗传变异程度较福建群体的高。有学者[30]也对广西北海、广东深圳、福建厦门三个黄鳍棘鲷的自然群体进行了遗传多样性分析,结果表明其自然群体存在丰富的DNA 序列遗传多样性。此外,还有学者[10,15]仅仅开发了一些黄鳍棘鲷微卫星标记,可为其遗传资源研究提供一定的基础资料。综上所述,中国南海区域的黄鳍棘鲷自然群体的遗传多样性程度较高。
该研究除了检测广东阳江自然群体的等位基因数,多态信息含量和杂合度之外,还分析了近交系数F、FIS等,这些指标可以反映种群适应力的遗传学指标。从近交程度中的近交系数F均值和FIS指标来看,目前广东阳江黄鳍棘鲷自然群体没有出现遗传衰退的现象,遗传适应能力较强。有研究表明近交系数均值小于0.1,生物种群才能避免受到近交抑制的影响[31]。而该种群F均值为0.034 2,证明广东阳江黄鳍棘鲷自然群体并没有受到近交抑制影响。
3.3 黄鳍棘鲷放流苗种遗传质量评估
基于6 个遗传参数,该研究比较了黄鳍棘鲷自然群体和放流苗种的遗传特征(表3)。结果表明除等位基因数NA外,放流苗种遗传多样性水平中的多态信息含量PIC、观测杂合度HO和期望杂合度HE均低于自然群体,此外自然群体近交指标中的近交系数F和FIS也低于放流群体,表明放流苗种在多方面存在遗传质量缺陷。
影响水生生物放流苗种遗传质量的因素主要有三点:亲本来源、数量和雌雄比例等[32]。该研究中的黄鳍棘鲷亲本来源于前期捕捞收集的广东阳江等海域的野生个体,表明了亲本个体的优良性。其次苗种亲本随机挑选于野生群体中,保证了亲本雌雄比例的合理性。此外,该研究收集的苗种393尾来自约100 尾亲本。Tajima 等[33]研究表明群体中个体数达到其有效种群大小的4~ 10 倍左右,才能促使该群体的遗传信息一代一代稳定传递。在针对红鳍笛鲷增殖放流苗种评估的研究中,付亚男等[6]指出造成苗种遗传质量偏低原因可能是与“亲本数量偏低”有关。而该研究中,黄鳍棘鲷的亲本数量仅为112 尾,远低于放流苗种亲本的合理数量,笔者推测黄鳍棘鲷放流苗种遗传质量缺陷可能是由亲本数量不足导致的。亲本的数量不足,会导致无法将完整的遗传信息传给下一代,造成子代遗传信息不足。
