海南东部外海海域鱼类群落结构季节变动及其影响因素
2021-06-12陆丽仪曾嘉维王学锋
陆丽仪,曾嘉维,2,林 坤,张 静,2,王学锋,2
(1.广东海洋大学水产学院,广东 湛江 524088; 2.南方海洋科学与工程广东省实验室(湛江),广东 湛江 524025)
鱼类是渔业资源数量最大的类群[1],其群落结构的时空分布与海洋环境密切相关,不同环境条件下鱼类群落组成和结构变化可作为评估海洋生态健康状况的指标,也是渔业资源的可持续利用与现代化管理的基础[2]。
南海近海渔业资源衰退严重,而外海海域渔业资源仍有一定的开发潜力[3]。目前,南海95%以上的捕捞强度集中于北部近海,较大的捕捞压力使渔获量不断下降,优质鱼类减少[4]。南海鱼类群落结构分析主要集中于珠江口[5]、北部湾[2]、大亚湾[6]等近海海域,而外海鱼类资源量、群落结构时空变化及其与环境因子关系等有待深入研究。海南岛东部外海海域位于南海西北部,属粤西和海南东部渔场,其鱼类资源丰富,是短尾大眼鲷(Priacanthus macracanthus)[7]、蓝圆鲹 (Decapterus maruadsi)[8]、深水金线鱼(Nemipterus bathybius)[9]等重要经济鱼类的产卵场。迄今,在南海西北部海域开展的空间大尺度鱼类群落研究较多[10-12],而海南外海海域等局部尺度上鱼类群落的调查尚未见报道。笔者研究该海域主要鱼类组成、资源量及共有优势种特征,运用冗余分析 (Redundancy analysis,RDA) 量化主要环境因子对群落结构的影响程度,为该海域的渔业资源变动及可持续利用研究提供参考。
1 材料与方法
1.1 数据来源
于2019 年4 月 (春季) 和9 月 (秋季) 对水深86~ 380 m 的海南东部外海海域进行渔业资源底拖网调查,共设置12 个站位 (图1)。调查船长29.6 m,宽6.4 m,主机功率368 kW。底拖网网囊网目为2.5 cm,上纲长度44 m。因每个站位的海况不同,实际拖速与时间可能有所偏差,数据处理时统一标准化为拖速3 kn,拖网时间1 h。渔获物的采集和分析按照《海洋渔业资源调查规范》[13]进行,利用YSI 水质检测仪测定溶解氧、pH、盐度和水温。

图1 2019 年海南东部外海海域采样站位Fig.1 Sampling stations of fishery resources in the off-sea of eastern Hainan in 2019
1.2 数据处理
1.2.1鱼类资源密度的估算 采用底拖网扫海面积法估算鱼类质量密度(D,kg/km2)和数量密度[2]。

式中,C为平均每小时拖网渔获量 (kg/h);a为每小时网具扫海面积 (km2/h);q为逃逸率,取值范围为0~ 1,本研究取0.5。
1.2.2优势度 使用相对重要性指数(IRI)[14-15]衡量各渔获物群落的生态优势度。

式中,N为某一种类的数量占总渔获数量的百分比(%);W为该种类的质量占总渔获质量的百分比(%);F为该种类出现的站位数占总站位数的百分比(%)。将IRI ≥1 000 的种类称为优势种,将1 000 >IRI ≥100 的种类称为重要种。
1.2.3体长频率分析 对共有优势种进行体长频率分析,利用Sturges 法和Snedecor 法分别计算分组组距,并求平均值作为分组参考[16],按照《海洋渔业资源调查规范》[13]及常用划分习惯确定组距。本研究暂定出现频率为10%以上的体长组定义为优势体长组。
1.2.4多样性指数计算 采用Margalef物种丰富度指数 (D)、Shannon-Wiener 多样性指数 (H′) 和Pielou 均匀度指数 (J′) 分析群落的物种多样性,并与近海海域各站位多样性指数H′ 进行单因素方差分析。由于鱼类个体大小差异较大,Wilhm[17]提出用质量代替数量计算H′ 值更为合理,故本研究根据质量密度计算群落物种多样性[18]。

式中,s为物种数;M为渔获鱼类总质量密度;Pi为第i种鱼类占总质量密度的比例,即
1.2.5多元统计分析 统计分析前,先对原始数据进行预处理。将出现频率小于10%的种类剔除,以减少稀有种对群落结构分析的影响[14]。同时,对各站位鱼类质量密度进行4 次方根转换,对除pH 外的环境因子进行lg (x+1) 转换[19]。基于鱼类群落的去趋势分析 (DCA) 结果,判断是用单峰模型(CCA) 还是用线性模型 (RDA) 以分析鱼类群落与环境因子的关系。若4 个轴中最大值 > 4,选用CCA 分析;若最大值< 3,则选用RDA 分析;若最大值在3~ 4 之间,则用RDA 和CCA 分析均可[20]。运用蒙特卡洛置换检验 (Monte Carlo permutation test) 分析鱼类群落分布与环境因子间的显著性[21]。以上数据分析用R 语言统计软件中vegan 包完成[22],站位图用Arcgis10.2 绘制。
2 结果与分析
2.1 种类组成
海南东部外海海域两个季度共捕获鱼类312 种,其中306 种鉴定到种,6 种鉴定到属,隶属于26 目96 科195 属。全部渔获中以鲈形目最多,共87 种(占种类总数的27.9%),其次为鲉形目和鲽形目,分别为50 种 (16.0%) 和35 种 (11.2%)。春季225种,秋季207 种,两个季度共有种数量为120 种,其中鲈形目45 种,鲉形目14 种,鲽形目和鳗鲡目10 种,其余目种类数均少于7 种。
2.2 优势种
春、秋季相对重要性指数分析结果见表1,主要物种均为底栖和中上层小型鱼类,两季共有优势种为黄鳍马面鲀 (Thamnaconus hypargyreus) 和深水金线鱼。春季优势种为黄鳍马面鲀和深水金线鱼,共占春季渔获数量的34.32%、渔获质量的34.88%,重要种15 种。秋季优势种则为蓝圆鲹、黄鳍马面鲀、深水金线鱼和短尾大眼鲷,共占秋季渔获数量的52.31%、渔获质量的58.09%,重要种13 种。

表1 海南东部外海海域鱼类群落主要物种相对重要性指数Table 1 Indices of relative importance (IRI) of fish species in the off-sea of eastern Hainan

续表1(Continued)
共有优势种中,黄鳍马面鲀秋季优势体长组比春季大,深水金线鱼秋季优势体长组比春季小,但体长范围比春季大 (图2)。黄鳍马面鲀春季体长范围为27~ 110 mm,优势体长组为60~ 85 mm (占总测定数62.3%);秋季体长范围为54~ 128 mm,优势体长组为80~ 105 mm (占70.1%)。深水金线鱼春季体长范围为48~ 155 mm,优势体长组为80~130mm (占88.0%);秋季体长范围为42~ 255 mm,优势体长组为90~ 120 mm (占57.8%)。秋季深水金线鱼20~ 70 mm 出现小高峰,渔获组成变化可能受伏季休渔和季节变化等方面的共同影响。

图2 春、秋季共有优势种体长频率分布Fig.2 Length-frequency analysis of common dominant species in spring and autumn
2.3 渔获量分布
个体平均质量 (平均质量密度/数量密度) 为秋季大于春季,表明秋季个体经伏季休渔和春夏生长,总体上体长增加,渔获物质量有所提升。平均质量密度秋季 (942.99 kg/km²) > 春季 (580.29 kg/km²),数量密度呈相反趋势,秋季 (29 283 尾/km²)< 春季 (52 362 尾/km²)。
主要物种渔获量的季节变动是导致各站位季节间渔获量变动幅度大的主要原因。春、秋质量密度和数量密度变化最大的是S5和S12站位 (图3)。S12 站位在春季中质量密度 (1 739.92 kg/km2) 及数量密度 (261 537 尾/km2) 最大,秋季质量密度和数量密度分别减少至210.33 kg/km2和22 598 尾/km2。S5 站位春季质量密度和数量密度分别仅为485.87 kg/km2和24 468 尾/km2,秋季质量密度 (5 311.45 kg/km2) 及数量密度 (116 237 尾/km2) 最大。

图3 海南东部外海海域质量密度和数量密度的空间分布Fig.3 Spatial distribution of fish biomass and abundance in the off-sea of eastern Hainan
2.4 物种多样性
秋季群落多样性水平 [Shannon-Wiener 多样性指数 (H′)、Pielou 均匀度指数 (J′) 和Margalef物种丰富度指数 (D)] 平均值均比春季略高,两个季节H′ 和J′ 值站位间变动均较大 (图4)。春季和秋季H′值分别为1.29~ 2.84 和1.88~ 3.09,平均值分别为2.18 和2.47。J′、H′ 值变化趋势大致相同,春、秋季变化范围分别为0.30~ 0.69 和0.46~0.75,平均值分别为0.55 和0.61。D值除S11 站位数值较大外,其余站位间波动不大,其春、秋季变化范围分别为4.03~ 7.42 和4.39~ 8.50,平均值分别为5.06 和5.67。

图4 海南东部海域鱼类群落各站位的多样性变化Fig.4 Diversity variations of fish communities at stations in the off-sea of eastern Hainan
2.5 鱼类群落时空分布与环境因子的关系
水深和pH 是影响春、秋两季鱼类群落结构的主要环境因子,盐度在秋季亦显著影响鱼类分布(图5)。春季水深 (P=0.003) 和pH (P=0.002) 对鱼类群落结构影响有高度统计意义 (表2)。5 个环境因子共解释物种数据 (种类组成和质量密度) 累计方差的58.34%,第1 排序轴与第2 排序轴分别解释29.73%和13.86%的物种分布变化。秋季水深 (P=0.004)、pH (P=0.017) 和盐度 (P=0.013) 对鱼类群落空间分布具统计意义,其中水深的影响最显著。环境因子共解释物种数据分布总变化的53.94%。第1排序轴与第2排序轴分别解释18.76%和13.06%的物种分布变化。

图5 海南东部外海海域鱼群落结构与环境因子的RDA 排序Fig.5 RDA triplots of fish community and environmental factors in the offshore waters of eastern Hainan

表2 海南东部外海海域环境因子相关显著性检验Table 2 Significance test of environmental factors in the eastern waters of Hainan
3 讨论
3.1 鱼类群落结构特征
3.1.1鱼类资源量状况 海南东部外海海域鱼类物种较丰富,春、秋两季总种类数 (312 种) 比同纬度北部湾近海海域 (68 种)[2]多78.2%,这可能是由该海域复杂的水文环境及地形造成的。海南东部外海海域受南海暖流、东南沿岸流等海流相互作用,海底地形及鱼类栖息环境比北部湾复杂,因此生物种类更多[10]。
受鱼类季节洄游的影响,本研究海域鱼类优势种和渔获量季节更替明显。如蓝圆鲹、长体圆鲹和红尾圆鲹等鲹科鱼类,春季从外海向近海海域作产卵洄游,在夏季航次中仅为常见种或稀有种;秋季由浅到深向外海洄游,成为秋季调查中主要物种[23]。整体上,秋季质量密度较春季高,数量密度较春季低。从两季主要物种渔获量和共有优势种体长分布频率推测,经过3 个多月的伏季休渔,鱼类幼体繁殖生长,使物种资源量增加。然而随着休渔期结束,渔民开始高强度捕捞,导致秋季数量密度短时间内迅速下降[24-25]。与南海部分近海海域比较:海南近海海域[26]及大亚湾海域[27]均主要以鲾科(Leiognathidae)、天竺鲷科(Apogonidae)等浅水鱼类为主要优势种,海南东部外海海域主要以黄鳍马面鲀、深水金线鱼和蓝圆鲹等为优势种;海南东部外海海域鱼类资源密度远高于大亚湾[28]和北部湾海域[28]。
3.1.2鱼类群落物种多样性 多样性指数是评价群落结构和功能的重要指标[29]。Shannon-Wiener 多样性指数 (H′) 与种类数和物种间个体分配是否均匀有关,而Margalrf 丰富度指数 (D) 则受种类数和质量密度影响[30]。本研究海域春季种类数虽比秋季多,但秋季多样性指数 (H′)、丰富度指数 (D) 和均匀度指数 (J′)平均值均高于春季,主要原因是秋季质量密度较春季高,且秋季物种间分配较均匀。站位间多样性指数存在明显变化,表明海南东部外海海域鱼类群落存在一定的空间异质性[30]。
比较本研究海域与雷州湾、大亚湾海域春秋两季多样性结果 (表3),均匀度指数 (J′)在三个海域中相差不大,表明均匀度受鱼类生境变化的影响较小,与前人的研究结果相符[31]。而对于丰富度指数(D) 和多样性指数 (H′),即使雷州湾和大亚湾海域调查调查站位数更多或使用的网目更小,均呈现出海南东部外海海域、雷州湾、大亚湾依次增加的趋势,符合大陆架海域多样性指数随水深增加而增加的规律[32]。已有研究表明,群落多样性与人类活动的干扰也有较大联系[33-34]。近岸海域由于过度捕捞、工厂废水和生活污水排放、海岸工程建设等影响,鱼类生活环境遭到破坏[34];而外海海域受人类活动的影响较小,鱼类资源量较丰富,多样性相对较高。
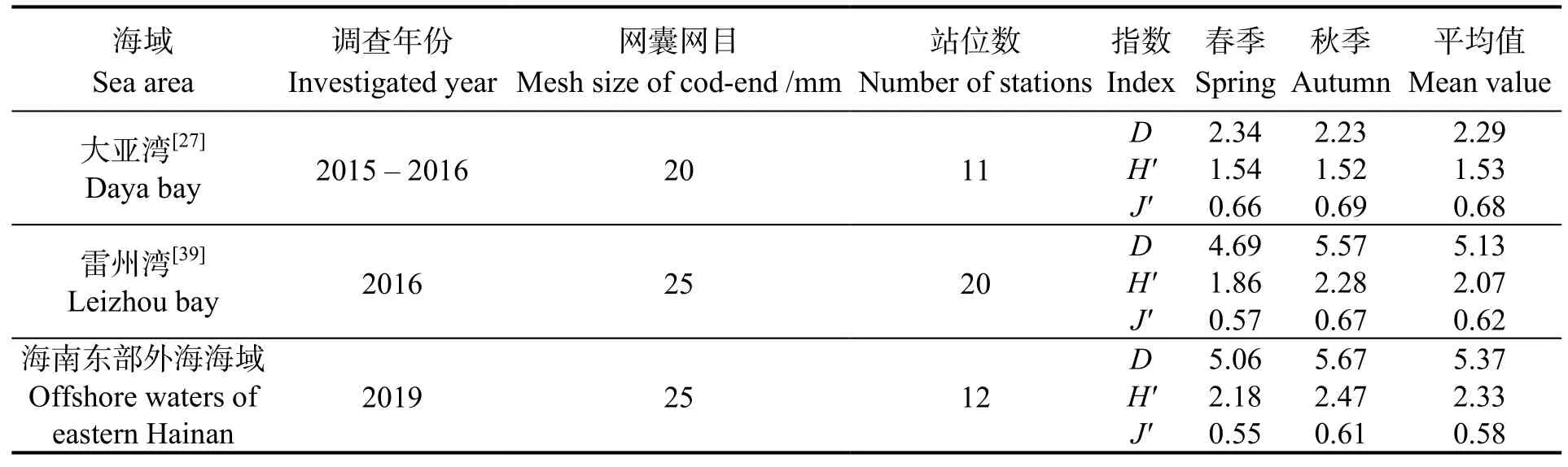
表3 海南东部外海海域鱼类群落与南海近岸海域比较Table 3 Comparison of fish community of offshore waters of eastern Hainan to the coastal waters of South China Sea
3.2 鱼类群落空间分布与环境因子的关系
RDA 分析不是针对某一变量进行分析,而是整体上获取能最大程度代表所有指标解释能力的最小变量组合[14]。其中,盐度仅在秋季对鱼类群落分布有重要影响,但实测数据中春季 (盐度33.69~34.64) 和秋季 (33.86~ 34.49) 的盐度变化均不明显,可能是因为春季中其他环境因子对群落分布更为显著而削弱了盐度对鱼类群落分布的影响[35]。pH作为综合性化学指标,是其他因素共同作用的结果,在一定空间范围内会受到盐度的影响[36]。本研究RDA 结果亦显示,pH 和盐度在春、秋两季中均呈正相关关系。另一方面,以上两种环境因素也会随着水深变化而发生相应的变化。南海北部大陆架研究表明,引起鱼类物种分布发生改变的不是水深本身,而是与水深有伴随关系的pH、盐度及光照等环境因素[37]。新西兰南部海域的鱼类群落在不同深度有不同的聚集模式[38]。本研究鱼类群落分布受水深影响显著的结论,与更大尺度范围的南海大陆架鱼类群落结构调查结果一致[37]。水深也是影响雷州湾[39]、大亚湾[27]等南海近岸海域鱼类群落分布的关键因子,即无论在近岸还是外海海域,水深均对鱼类群落结构有重要影响。
综上,本研究认为在海南东部外海海域的渔业资源量化管理中,除应关注主要经济种类种群水平上的捕捞产量外,更应注重群落水平的多样性变化、优势种的生物量及其变化因素。
