2018 年长江安庆段洄游汛期刀鲚年龄结构和生长特征
2021-06-12罗宇婷刘思磊马凤娇
罗宇婷,代 培,刘思磊,马凤娇,尤 洋,,刘 凯,
(1.南京农业大学无锡渔业学院,江苏 无锡 214081;2.农业农村部长江下游渔业资源环境科学观测实验站,中国水产科学研究院淡水渔业研究中心,江苏 无锡 214081)
刀鲚(Coilia nasus),属于鲱形目 (Clupeiformes)鳀科 (Engraulidae) 鲚属 (Coilia)[1],又名长颌鲚,俗称刀鱼,是一种典型溯河洄游性鱼类,也是长江中下游最名贵鱼类之一,与鲥 (Tenualosa reevesii)、暗纹东方鲀 (Takifugu obscurus) 合称为“长江三鲜”[2]。每年初春季节,长江刀鲚聚集于长江口近海区,集群沿江逆流而上到达产卵场完成繁殖活动,随后亲本返回近海育肥越冬[3]。
鱼类年龄结构和生长特征是渔业资源研究中的重要内容,可为研究鱼类个体的生长、种群的存活率、性成熟年龄及繁殖提供基础数据,有助于了解种群结构特征、衡量种群的生长状况[4],同时可为渔业主管部门的科学管理提供依据[5]。如孙雪兴等[6]于 1987 年通过对太湖湖鲚(Coilia ectenes taihuensis)年龄与生长的研究得出其合理的捕捞规格;周成等[7]于2012 年对印度洋南部大眼金枪鱼(Thunnus obesus)年龄与生长的研究得出以鳍条作为其年龄鉴定材料最适的切割位置;江艳娥等[8]于2019 年研究南海北部陆架水域多齿蛇鲻(Saurida undosquamis)与花斑蛇鲻(Saurida tumbil)的年龄与生长,发现其渔业资源衰竭,建议降低南海近海捕捞压力。Von Bertalanffy 生长方程是研究鱼类生长的适宜模型之一,可准确描述鱼类随年龄的生长规律,比较不同地理的种群生长特征[9]。已有学者采用 von Bertalanffy 生长方程描述麦穗鱼(Pseudorasbora parva)[10]、泉 水 鱼(Pseudogyrincheilus procheilus)[11]、裸腹叶须鱼(Ptychobarbus kaznakovi)[12]鱼类的生长特征。
现有的刀鲚研究资料主要集中于洄游特性[3,13]、遗传学[14]、繁殖学[15]及耳石指纹元素特征[16]等方向,对其年龄研究包括长江口[17]、长江无为江段[18]、九段沙、靖江、芜湖江段[19]及池养刀鲚的生长[20],针对刀鲚洄游汛期内全汛期连续采集样本进行的相关研究尚未见报道。本研究在洄游汛期内针对长江安庆段刀鲚进行连续采样,较为系统地研究样本群体年龄结构与生长特征,以期掌握该时段内长江安庆段刀鲚种群的年龄及生长特征,为刀鲚资源保护积累研究素材。
1 材料与方法
1.1 样本采集与处理
2018 年4-7 月于长江安庆段设置4 个样区利用流刺网采集刀鲚样本(图1),具体采样时间分布在4 月17 d,5 月21 d,6 月25 d,7 月21 d,选取双日刀鲚样本作为本次研究对象。网具由2 层衬网和1 层内网组成,网长为200 m,网高为2.8 m,衬网网目尺寸为28 cm,内网网目尺寸为3 cm。调查期间共采集刀鲚样本392 尾,现场对样本进行编号,测量体长(L,cm)、体质量(m,g)等生物学信息(体长精确到0.01 cm,体质量精确到0.01 g)。取15~ 20枚鳞片作为年龄鉴定材料,选取的鳞片需形态规则且位于鱼体中段侧线上方到背鳍下方(图2)。
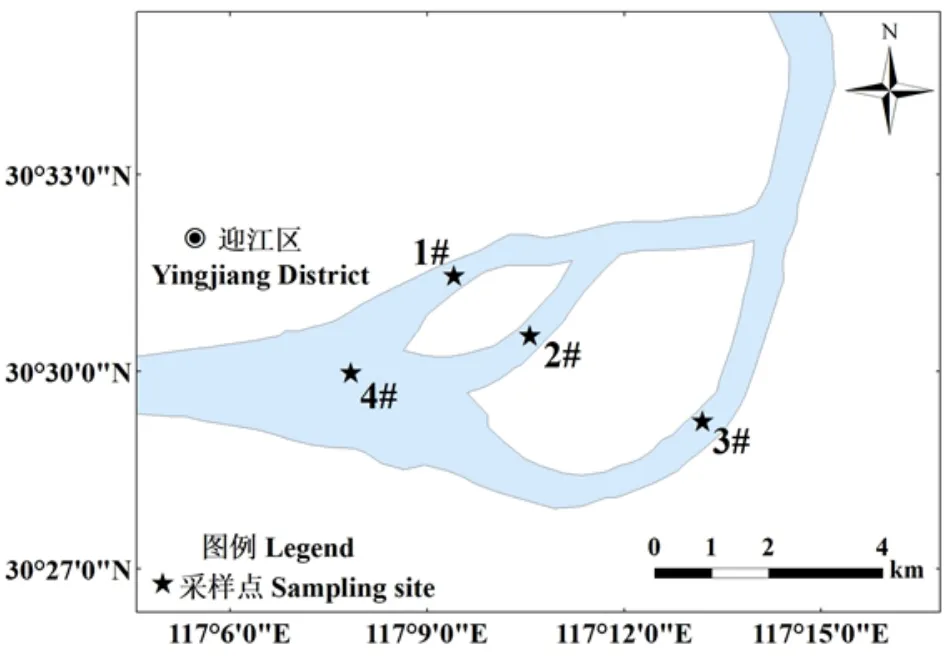
图1 刀鲚采样点位Fig.1 Sampling sites of Coilia nasus

图2 鳞片采集区域Fig.2 Scale collection area
1.2 鳞片处理与年龄鉴定
将鳞片放置于体积分数为5%氨水中浸泡24 h,待表层上皮组织和色素细胞降解后,剔除形状不规则的鳞片和再生鳞,用棉签、纸巾等洗净擦干并选择3~ 5 片夹片制作成标本并做好标记,在Nikon Zoom 645s 生物解剖镜下观察鉴定(图3)。

图3 刀鲚典型鳞片Fig.3 Typical scales of Coilia nasus
开展年龄鉴定时,在不依赖于其他生物学信息的基础上,作出第一次年龄鉴定;全部完成后,不以第一次的测定结果为依据的前提下再进行第二次鉴定;对比两次鉴定结果,对两次鉴定结果不一致的样本个体进行第三次鉴定。若第三次鉴定结果与前两次都不一样,则舍弃这一样本。
测量鳞片鳞焦沿辐射沟的直线长度与侧区两侧长度(图3 红线部分),三者均值作为鳞片鳞径(R,mm),同样的方式测得各年轮的轮径(图3黄线部分)。
1.3 数据统计与分析
数据记录:以冬龄记录,1 冬龄简写为1 龄,2 冬龄简写为2 龄,以此类推。
数据处理:
(1)体长与体质量关系式:

式中,m为实测体质量(g),L为实测体长(cm),b为幂函数指数。
t检验法[21]检验体长与体质量回归方程的幂指数“b”与“3”之间的差异:
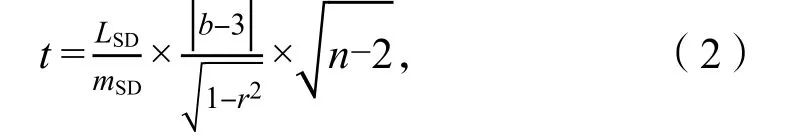
式中,LSD、mSD分别为体长、体质量对数值的标准差,n为标本数,r为相关系数
(2)体长与鳞径关系式:

式中,R为鳞径。
(3)Beverton 法和Ford 方程[22]:
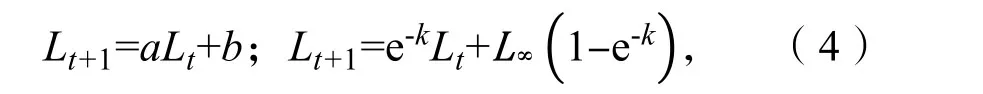
式中:Lt和Lt+1分别代表t龄和t+1 龄的体长(cm),k为生长系数
(4)年龄与生长关系用von Bertalanffy 方程[23]描述:

式中,L∞、m∞分别指渐进体长、渐进体质量,k为生长系数,t为年龄,t0为初始生长年龄。
(5)生长拐点方程[24]:

(6)对体长、体质量生长方程一次微分、二次微分得出体长、体质量的生长速度和生长加速度方程:
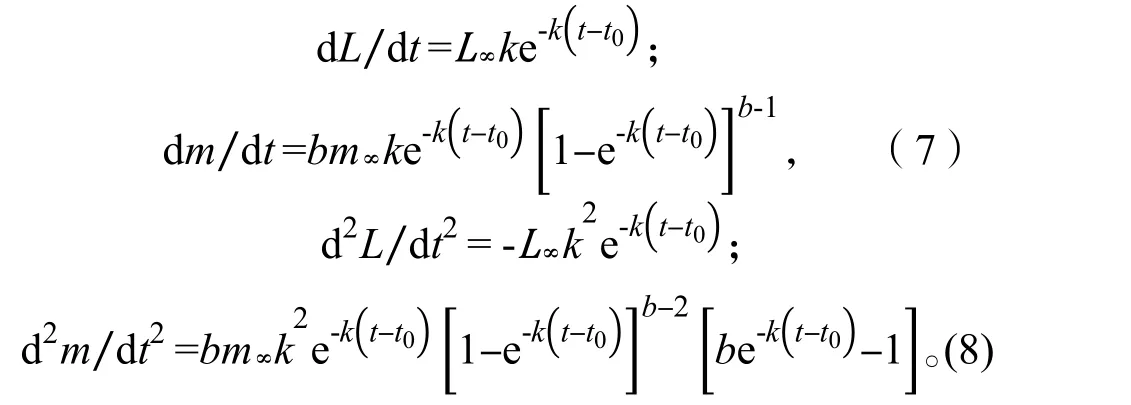
(7)生长指标方程[25]:
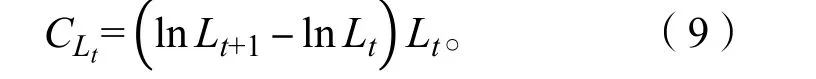
以上数据统计分析及图形绘制采用Microsoft Excel 2003 和 SPSS 20.0 进行。
2 结果与分析
2.1 渔获规格
本研究共采集392 尾长江刀鲚样本,其体长范围为18.99~ 32.00 cm,均值为(25.09 ± 2.05)cm,优势体长组为22.00~ 28.00 cm(图4),占样本总数的86.48%。体质量范围为18.33~ 119.67 g,均值为(51.43 ± 15.61)g,优势体质量组为(30.00~ 60.00)g(图5),占样本总数的70.40%。

图4 刀鲚体长的分布Fig.4 Distribution of body length of Coilia nasus
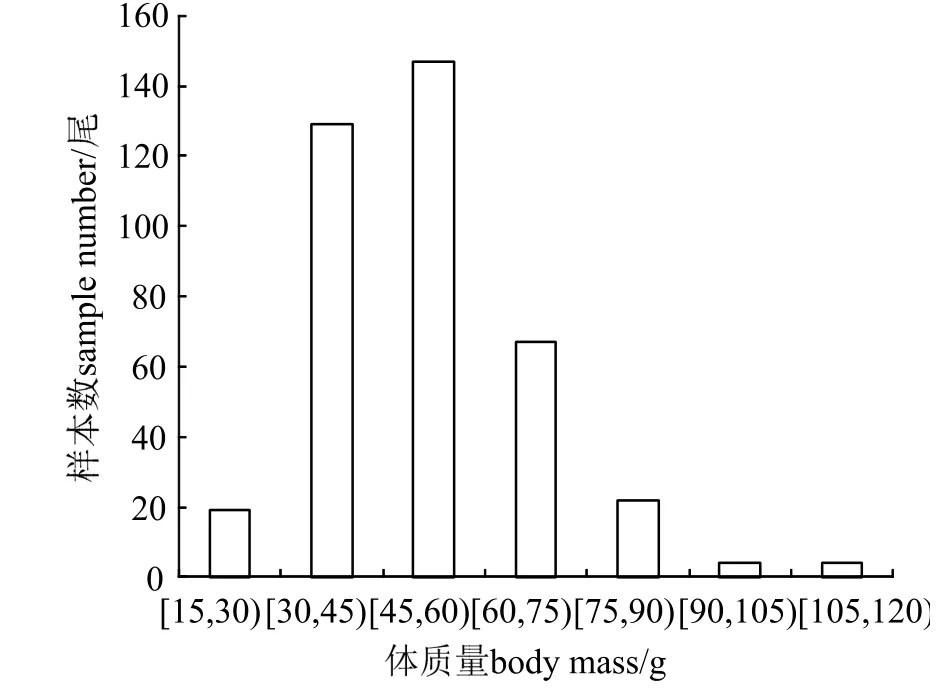
图5 刀鲚体质量的分布Fig.5 Distribution of body weight of Coilia nasus

图6 刀鲚体长与体质量的关系Fig.6 The relation between body length and body weight of Coilia nasus
2.2 年龄结构
刀鲚样本年龄组成为1~ 4 龄,其中优势年龄组为2 龄,占样本总数的47.96%;其次为1 龄,占40.82%;3 龄个体数较少,占比为10.97%,4 龄个体仅发现1 尾。刀鲚优势年龄组时间特征表现:4、5 月份优势年龄组为1 龄,分别占当月样本总数的83.33%、80.41%;6、7 月优势年龄组为2 龄,占比分别为60.59%、83.93%(表1)。

表1 刀鲚样本不同月份的年龄结构分布Table 1 Age structure of different month of Coilia nasus
2.3 生长特征
2.3.1体长与体质量的关系 幂函数拟合刀鲚体长与体质量关系回归方程为:m=0.001 3L3.2866(R2=0.854 6) (图6);回归方程中的幂函数“b”值与“3”之间差异不显著(t=1.94,P >0.05)。
2.3.2体长与鳞径的关系 体长与鳞径关系(图7)拟合方程为L=4.250 3R+14.008 (R2=0.467 6)。
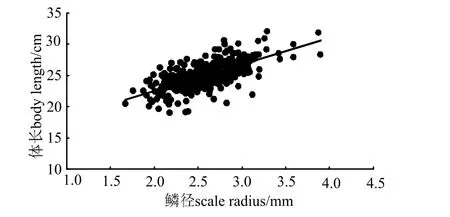
图7 刀鲚体长与鳞径的关系Fig.7 Regression linear for body length and scale radius of Coilia nasus
根据鳞径逆算各龄长江刀鲚样本的体长(表2)并与实测平均体长进行配对检验,结果表明逆算体长和实测体长间无差异(P >0.05)。
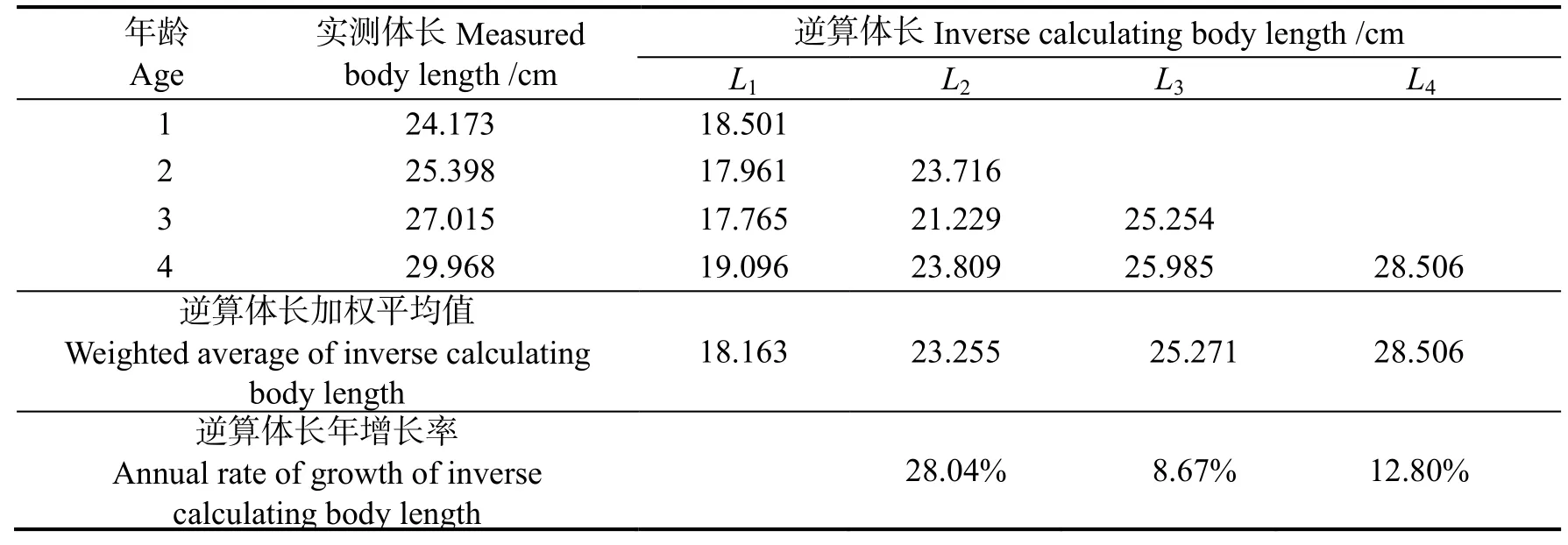
表2 刀鲚样本的逆算体长Table 2 Inverse calculating body length of Coilia nasus
2.3.3生长参数和生长方程 选用von Bertalanffy生长方程来拟合刀鲚体长和体质量的增长,通过Beverton 法和Ford 方程求得:L∞=32.75 cm,k=0.397 3,t0=-0.674 3;由体长和体质量关系式求得m∞=124.19 g。将上述参数代入von Bertalanffy 生长方程,得到刀鲚体长和体质量生长方程(图8):Lt=32.75[ 1 -exp (-0.397 3 (t+0.674 3)) ];mt=124.19[1-e-0.3973(t+0.6743)]3.2866。
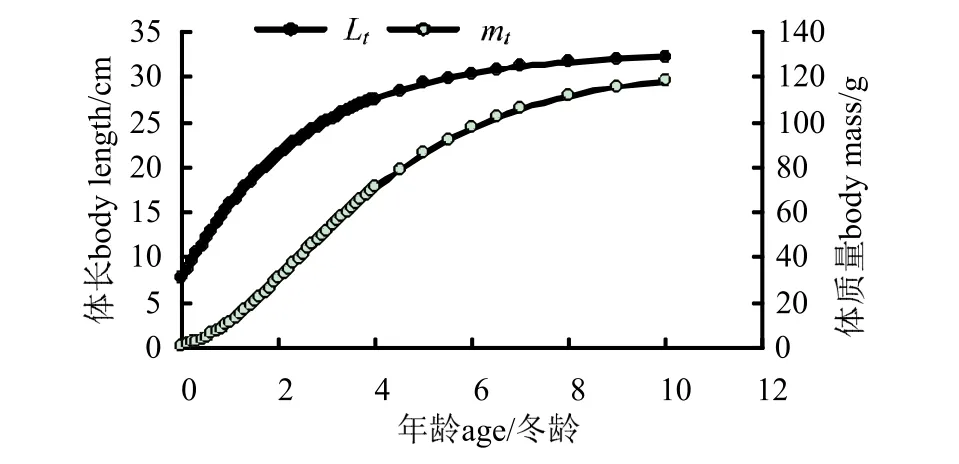
图8 刀鲚体长和体质量生长曲线Fig.8 The growth curve of body length and body weight of Coilia nasus
刀鲚体长和体质量均随着年龄增长而增加,其中体长生长是一条不具拐点的曲线,增长先快后慢,趋势随着年龄增加逐渐平缓,最终趋向于极限体长32.75 cm;体质量的生长曲线是具有拐点的S 型曲线,增长趋势先慢后快,再趋于平缓,最终趋向于极限体质量124.19 g。
2.3.4生长速度与生长加速度 体长和体质量生长曲线只能反映生长过程的总和,为了研究生长速度随时间的变化特征,分别对刀鲚体长、体质量生长方程一次微分、二次微分,得出体长、体质量的生长速度和生长加速度方程。
体长生长速度:
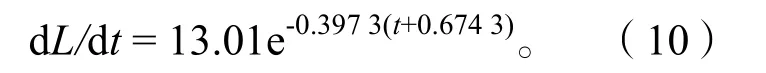
体长生长加速度:
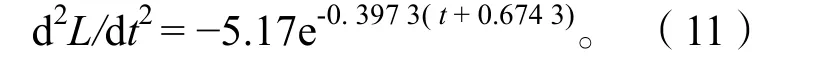
体质量生长速度:
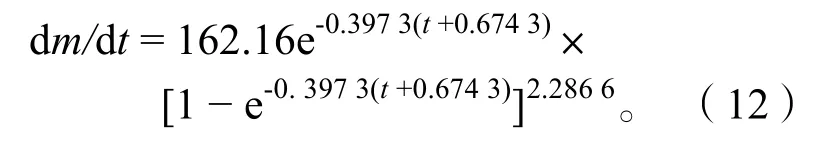
体质量生长加速度:
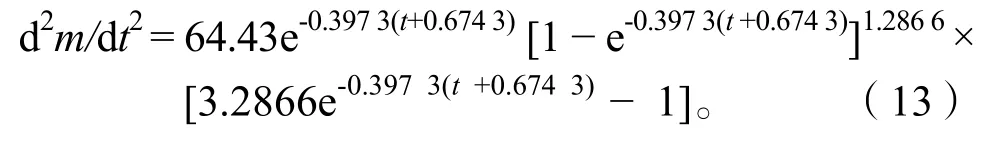
通过以上方程作出体长、体质量的生长速度曲线(图9)和生长加速度曲线(图10)。长江安庆段刀鲚生长拐点年龄为:ti=t0+lnb/k=2.32,对应体长Li=22.79 cm、体质量mi=37.69 g。

图9 刀鲚体长和体质量生长速度曲线Fig.9 The growth rate curve of body length and body weight of Coilia nasus
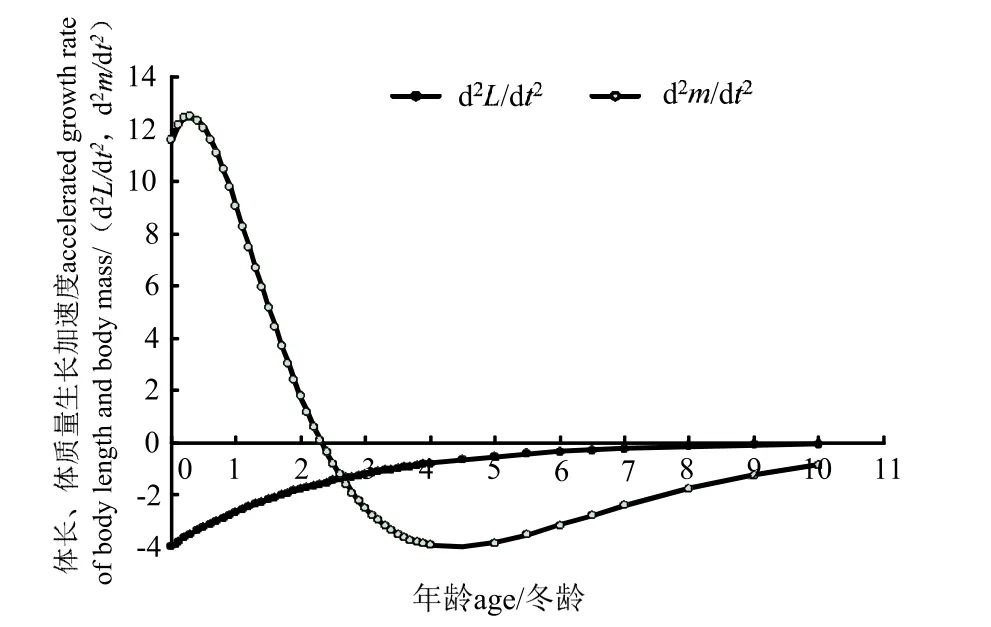
图10 刀鲚体长和体质量生长加速度曲线Fig.10 Accelerated growth rate curve of body length and body weight of Coilia nasus
长江安庆段刀鲚的体长生长速度和加速度曲线均不具拐点,其中生长速度为正值,随年龄增加而减少,减少趋势先快后慢逐渐平缓,最后趋向于零。生长加速度为负值,生长速度递减,减少趋势先快后慢,逐渐平缓并趋向于零。
长江安庆段刀鲚的体质量生长速度均为正值,体质量始终处于增长中。曲线具有一个拐点ti=2.32,即t< 2.32 龄时,体质量持续增长,增长速度逐渐加快,但加快的速度逐渐变慢;到t=2.32 龄时生长加速度为零,此时体质量生长速度达到最大;待t> 2.32 龄时,体质量生长速度逐渐下降,下降的速度先快后慢,最后趋向于零。
2.3.5生长指标 生长指标主要用于比较鱼体的生长速率,根据逆算体长得出长江刀鲚体长的增长率与生长指标(表3)。由于4 龄鱼仅1 尾,不能客观代表该年龄组的生长情况,不具参考价值。可以看出1~ 3 龄鱼始终保持生长趋势,短期内体长增长率和生长速率随年龄增加而增长。
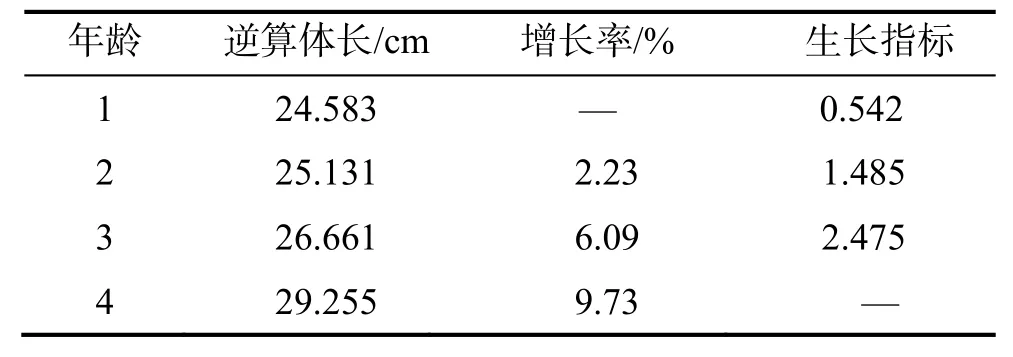
表3 刀鲚体长的生长指标Table 3 The growth index of body length of Coilia nasus
3 讨论
3.1 长江安庆段刀鲚的生长特征
鱼类年龄鉴定方法主要包括硬组织鉴定法和长度分析法,其中硬组织鉴定法得到了广泛应用[26],可用于鱼类年龄鉴定的硬组织包括鳞片、耳石、脊椎骨、鳃盖骨、鳍条等。1898 年鳞片第一次被用于鱼类年龄鉴定[27],由于其相比其他材料采集处理方便且易于识别年龄,因此一直是鱼类年龄鉴定的重要材料。
鱼类生长过程中体长和体质量的关系,是鱼类基础生物学研究重要部分,对渔业捕捞生产具有指导价值。本研究中刀鲚体长与体质量的幂函数方程为m=0.001 3L3.2866,其中b值为3.286 6,经t检验与“3”无差异,表明长江安庆段刀鲚群体在体长、体高、体宽三个方向生长速度大致相等[28]。b反映鱼类在不同阶段环境中生长情况,本研究b值略低于2006 年长江口崇明岛刀鲚群体 (b=3.36)[17]、2006 年长江无为段刀鲚群体 (b=3.331 2)[18]及池养刀鲚(b=3.592 4)[20],高于不同研究期(2003-2004,2008-2009,2012-2013)靖江段刀鲚群体(b=2.961~ 3.061,b=3.086 6)[29-30],这表明不同水域刀鲚群体的生长速度存在差异,这可能与研究期、样本量和水文条件有关,具体原因有待进一步研究。
本研究推算的刀鲚极限体长、体质量分别为32.75 cm,124.19 g,生长参数k=0.397 3,满足e-k(0.672 1)<1 的理论,表明von Bertalanffy 方程能较好拟合生长[31]。郑飞等[19]对2009 年采自长江九段沙、靖江和芜湖3 个江段的299 尾洄游型刀鲚样本的调查结果显示最大体长、体质量分别为32.8 cm、143.80 g,略高于本次研究结果;吴斌等[32]对2013年至2014 年鄱阳湖刀鲚调查显示极限体长体质量分别为40.95 cm 和230.09 g,也高于本研究安庆段刀鲚的极限体长体质量数据。
董文霞等[29]研究的2012 年长江靖江段刀鲚鲚群体的初始生长年龄t0为-0.55 龄,拐点年龄ti为2.99 龄;郭弘艺等[30]研究的不同时期(2003-2004,2008-2009,2012-2013)靖江段沿岸刀鲚群体t0为-0.49 和-0.50 龄,ti为1.49~ 1.54 龄;吴斌等[32]研究2013-2014 年鄱阳湖刀鲚群体t0为-0.57 龄,拐点年龄为4.00 龄。本研究显示,长江安庆段刀鲚群体初始生长年龄为-0.67 龄,体质量生长速度拐点年龄为2.32 龄,与之前的研究相比初始生长年龄相对较早但相差不大,而拐点年龄差别较大,刀鲚最小1 冬龄便可达到初次性成熟[29](1 龄样本占40.82%),而生长拐点的到来在性成熟期之后,表明部分刀鲚在达到初次性成熟后仍会经过一段快速生长期,这与本研究生长指标的数据结果相符合。本研究生长参数k值为0.397 3,介于0.20~ 0.50 之间,属于快速生长鱼类[33],但与2006 年崇明岛附近长江口水域的刀鲚群体(k=0.51)[17]和2010-2011 年淀山湖水域刀鲚(定居型,又称湖鲚)群体的k值(0.54)[34]相比有所下降,与2012 年长江靖江段刀鲚群体(k=0.38)[29]相比略微上升,一方面体现了该江段刀鲚洄游群体的生物学特征,另一方面也表明刀鲚呈现出小型化趋势。
3.2 长江安庆段刀鲚种群年龄结构
20 世纪70 年代初,长江刀鲚群体年龄由0~ 6龄组成,且3、4 龄占年龄组成七成以上[19],表明进行多次洄游产卵的剩余群体占据优势;郑飞等[19]的调查显示2009 年长江下游三个江段刀鲚的优势年龄组为2 龄,占全部个体的57.53%,其次为3龄,占40.80%,1 龄和4 龄个体均很少;张敏莹等[35]对1993-2002 年长江下游五个监测点刀鲚的调查显示汛期种群以2 冬龄为主。本实验刀鲚的年龄结构为1~ 4 龄,优势年龄组为1~ 2 龄(占88.78%),4 龄鱼仅发现1 尾,可见长江安庆段刀鲚群体结构以首次到达性成熟的补充群体或低龄的剩余群体占据绝对优势。鱼类生长规律是低龄鱼生长强度大[36],长江刀鲚也不例外,低龄群体生长发育快,体内脂肪大量累积,在汛期开始阶段可能优先进行洄游,因而实验结果显示4、5 月份中的1 龄鱼占多数;而2 龄或2 龄以上长江刀鲚可能洄游开始较晚,因而在渔汛期中后期个体较多。不仅如此,捕捞胁迫对鱼类群体的表型性状有着重要的影响,引起鱼群低龄化、小型化、性早熟等现象[37],对长江刀鲚的生殖洄游过程同样有影响。在长江刀鲚自长江口向长江洄游过程中,将历经重重捕捞,大规格的高龄个体到达长江安庆段的概率已经非常小,这与本实验调查结果相符,即种群结构以低龄的1 龄和2 龄占绝对优势。需要注意的是,据历史资料记载,长江刀鲚洄游群体以2 冬龄以上个体占优[19,38],随着捕捞压力的不断增大,长江刀鲚资源不断衰退,洄游群体趋于小型化和低龄化,本实验结果显示,1 龄的个体占比高达40.82%,这也充分反映了刀鲚低龄化的趋势。
近年来,水环境污染、涉水工程建设和过度捕捞等人类扰动对包括刀鲚资源在内的长江渔业资源造成了巨大胁迫,随着刀鲚资源不断衰退,捕捞量大幅下降,市场上刀鲚价格急剧攀升,高额利润驱使渔民增大捕捞强度,形成恶性循环[38]。在此背景下,渔业主管部门已于2019 年起全面停止长江刀鲚的商业捕捞,实施资源养护。结合禁捕措施,应进一步加强长江刀鲚洄游生态学及重要栖息地保护研究,系统地保护和恢复这一珍稀鱼类资源。
